Dreaming and the default network: A review, synthesis, and counterintuitive research proposal
G. William Domhoff
Department of Psychology, University of California, Santa Cruz
Kieran C. R. Fox
Department of Psychology, University of British Columbia
|
NOTE: If you use this paper in research, please use the following citation, as this on-line version is simply a reprint of the original article:
|
|
Domhoff, G. W., & Fox, K. R. (2015). Dreaming and the default network: A review, synthesis, and counterintuitive research proposal. Consciousness and Cognition, 33, 342-353.
|
Abstract
This article argues that the default network, augmented by secondary visual and sensorimotor cortices, is the likely neural correlate of dreaming. This hypothesis is based on a synthesis of work on dream content, the findings on the contents and neural correlates of mind-wandering, and the results from EEG and neuroimaging studies of REM sleep. Relying on studies in the 1970s that serendipitously discovered episodes of dreaming during waking mind-wandering, this article presents the seemingly counterintuitive hypothesis that the neural correlates for dreaming could be further specified in the process of carrying out EEG/fMRI studies of mind-wandering and default network activity. This hypothesis could be tested by asking participants for experiential reports during moments of differentially high levels of default network activation, as indicated by mixed EEG/fMRI criteria. Evidence from earlier EEG/fMRI studies of mind-wandering and from laboratory studies of dreaming during the sleep-onset process is used to support the argument.
|
Introduction and overview
The general outlines of the neural correlates of dreaming, which are putative indices of the neural substrate that supports dreaming, have been known since the late 1990s due to the unexpected convergence of findings from two very different types of studies: neuroimaging studies of REM sleep on the one hand, and neurological examinations and CT scans of patients with alterations in their dreaming on the other (e.g., Braun et al., 1997; Maquet et al., 1996; Nofzinger, Mintun, Wiseman, Kupfer, & Moore, 1997; Solms, 1997). The neuroimaging and neurological literatures were then synthesized with the literatures on dream content and on the gradual development of frequent and complex dreaming in children to create a neurocognitive theory of dreaming (Domhoff, 2001, 2003, Chapter 1). Around the same time, a serendipitous discovery showed that a "default" network of brain regions is spontaneously active during restful states, and supports mind-wandering and daydreaming during waking (e.g., Addis, Wong, & Schacter, 2007; Andrews-Hanna, Reidler, Huang, & Buckner, 2010; Christoff, Gordon, Smallwood, Smith, & Schooler, 2009; Dixon, Fox, & Christoff, 2014; Gusnard & Raichle, 2001; Kucyi & Davis, 2014; Mason et al., 2007). Not only is the default network activated during waking daydreaming; recent findings show that many hubs of the default network are even more active during REM sleep than at rest, augmented by secondary visual and sensorimotor cortices that support sensorimotor imagery (Chow et al., 2013; Fox, Nijeboer, Solomonova, Domhoff, & Christoff, 2013).
It is also possible that this augmented default network could support episodes of dreaming during the complex and multifaceted sleep-onset process and into Stage 2 of NREM, before Stages 3 and 4 occur (e.g., Horovitz, 2008; Larson-Prior et al., 2009; Laufs, 2008, for a review; Laufs, Walker, & Lund, 2007; Sämann et al., 2011; Tagliazucchi et al., 2013). Based on the similarity of dream reports after spontaneous morning awakenings from REM and NREM 2 (Cicogna, Natale, Occhionero, & Bosinelli, 1998), and the higher level of general brain activation in the early morning hours (Antrobus, Kondo, & Reinsel, 1995; Wamsley et al., 2007), it seems likely that the putative neural network for dreaming is active during NREM 2 at this time as well (Domhoff, 2011b, pp. 1168--1169, for a review of the evidence). However, due to the difficulties of investigating the full sleep cycle with noisy brain scanning equipment, no neuroimaging study has yet examined NREM 2 in the early morning.
Despite the progress in outlining a general pattern of brain activity during dreaming, further advances in specifying the neural correlates of dreaming may be slow, for at least two reasons. First, cognitive neuroscientists interested in the functioning of the default network are rightly focused on many other issues, including the role of this network in social-affective cognition and in various neurodegenerative, neurodevelopmental, and psychiatric conditions, such as Alzheimer's, autism, and schizophrenia (e.g., Amft, Bzdok, Laird, Fox, & Schilbach, 2014; Andrews-Hanna, 2012; Andrews-Hanna, Smallwood, & Spreng, 2014, for overviews). Second, the type of studies of the default network and its relationship to ongoing mentation that are needed (namely, combined EEG/fMRI studies) are more difficult to conduct during sleep because of the noise of the fMRI scanner, the fact that it takes an entire night to adapt to sleeping in a laboratory in the best of circumstances, and the relatively time-consuming process of awakening participants from sleep and having them attempt to recall often hazy mental experiences. Adding to the difficulty of studying dreaming during sleep or sleep onset in conjunction with EEG/fMRI, participants in these studies are usually sleep deprived or instructed to shorten their normal sleeping period for several days beforehand, which reduces functional connectivity in the default network and may alter the production and recall of dreams (De Havas, Parimal, Soon, & Chee, 2012).
Given these and other problems, it seems likely that new approaches compatible with the primary interests of neuroimaging researchers are needed for studies of the neural correlates of dreaming. It is the purpose of this article to suggest one such approach. This approach is counterintuitive because it involves studying dreaming during brief episodes of dreaming during mind-wandering in the waking state, a phenomenon that was accidentally discovered in the 1970s (Foulkes & Fleisher, 1975; Foulkes & Scott, 1973). More specifically, the necessary EEG/fMRI studies might be done as part of larger studies of the default network carried out for other purposes.
What is dreaming and when does it occur?
There is no consensus on the definition of dreaming. For example, dreaming is sometimes considered to be a type of hallucinatory activity, which is defined very generally as perceiving something that isn't there, or as the perception of sights and sounds with no reality basis. This view is often adopted in neurophysiological theories of dreaming, which leads to the idea that dreaming is a form of psychosis. For example, psychiatrist J. Allan Hobson (2002, pp. 98--99) asserts, "psychosis is, by definition, a mental state characterized by hallucinations and/or delusions," so it follows that dreaming is "as psychotic a state as we ever experience while awake." Similarly, the Freudian theorists Mark Solms and Oliver Turnbull (2002, p. 213) argue that "the functional anatomy of dreaming is almost identical to that of schizophrenic psychosis," which is characterized by frequent hallucinatory activity. On the other hand, dreaming sometimes is defined very broadly as any form of mental activity during sleep, which can range from thoughts and isolated images to the more story-like experiences that are recalled upon awakening; in still other studies, ordinal scales are used to rate mentation reports as to the degree that they are like canonical definitions of dreaming, which can bring about considerable agreement concerning the amount of dreaming that was found in a wide range of studies (e.g., Nielsen, 2000; Pagel et al., 2001; Zadra & Domhoff, 2011, for further discussion).
Based on analyses of thousands of reports from hundreds of children and adults that were studied in dream research laboratories, we emphasize the content of dream reports in defining dreaming as an involuntary but organized mental act that generates credible simulations of the real world (c.f. Foulkes, 1985, Chapter 1). This definition is consistent with the way in which simulation is defined by mind-wandering researchers, namely as "a particular kind or subset of thinking that involves imaginatively placing oneself in a hypothetical scenario and exploring possible outcomes" (Schacter, Addis, & Buckner, 2008, p. 42). This definition also decouples dreaming from sleep, making it possible to test the idea that episodes of dreaming can occur during periods of mind-wandering through the use of the same content indicators and rating scales that are used to differentiate the various types of reports that are collected from REM and NREM awakenings in sleep laboratories (Domhoff & Schneider, 1999, pp. 149--150; Foulkes, 1985, pp. 58--64; Hall & Van de Castle, 1966).
Moreover, this definition is consistent with the idea that the mental imagery during dreaming is best understood as "embodied simulation." The concept of embodied simulation as we use it is based on experimental evidence from psychological research, which reveals that brain areas supporting visual and sensorimotor processing are also activated when purely mental imagery is generated (in the absence of perceptual inputs); in addition, the imagery involved in simulation, whether literal or figurative, is subjectively "felt" as the experienced body in action (e.g., Bergen, 2012, pp. 13--17; Gibbs, 2006, 2014, pp. 27-- 28; Niedenthal, Winkielman, Mondillon, & Vermeulen, 2009). Then, too, literal and figurative meanings for the same word, in this instance the motion verb "run," lead to slightly different brain activations for fictive motion in an fMRI study (Lauro, Mattavelli, Papagno, & Tettamanti, 2013). The case for embodied simulation as essential to "off-line" cognition also has been made from studies of working memory, episodic memory, implicit memory, and even reasoning (Wilson, 2002, pp. 632--634, for a synthesis of work by herself and others).
Thus, dreaming can be understood as a form of embodied simulation in which a person's imaginings are more fully enacted and dramatized than during waking thought. As an enactment, dreaming includes the dreamer as an observer or participant, and also usually includes at least one other character as well (e.g., Domhoff, 1996; Hall & Van de Castle, 1966). In addition, the dreamer and the other characters in dreams are invariably engaged in one or another activity (e.g., looking, talking, walking, running) or a social interaction (e.g., befriending, arguing, flirting) within a familiar or unfamiliar setting. It is this sense of being an embodied participant in (or observer of) an event, along with the inclusion of characters, activities, and social interactions, that distinguishes dreaming from other forms of thinking, whether during sleep or waking (Domhoff, 2011a, 2011b).
This empirically derived characterization of dreaming also suggests that dreaming is very different from any form of waking hallucinatory experience. For example, a distinction between hallucinating and dreaming is made repeatedly by the neurologist Oliver Sacks (2013, pp. xiii, 26--27, 80, 209, 214) in his book-length discussion of the wide variety of hallucinations that have been reported in the literature, many of which are experienced by people without any psychiatric problems. Sacks's (2013) review of the literature, along with his own observations in the course of treating neurological patients, are further supported by reports from several hospitalized schizophrenics, who told the psychiatrists who interviewed them that their hallucinations were not like their dreams (Kass, Preiser, & Jenkins, 1970). In the case of psychiatric patients, their hallucinations are very frequently auditory, with persecutory figures interrupting their thinking or issuing commands, or involve highly repetitive visual imagery, neither of which resembles embodied simulations that include characters involved in activities and interactions that unfold in a constructed social world.
Besides its firm grounding in empirical research, another benefit of this definition, as briefly noted earlier in this section, is that it does not limit dreaming to any particular stage of sleep, nor even to sleep itself. Admittedly, much of the neuroimaging research that provides the basis for the working hypothesis presented here involves studies only of REM sleep (e.g., Braun et al., 1997; Maquet et al., 1996; Nofzinger et al., 1997, 1999) or of the overlap between REM and the waking default network (Fox et al., 2013, pp. 6, 11--12). Nonetheless, the similarities among dream reports from the sleep-onset process, NREM 2 late in the sleep period, and REM, along with preliminary evidence of brain activation at sleep onset and during NREM 2, leads to the hypothesis that animates this article: the same neural network that seems to support dreaming during REM is active at other times as well (Antrobus et al., 1995; Cicogna et al., 1998; Domhoff, 2011b; Foulkes, 1985; Ioannides, Kostopoulos, Liu, & Fenwick, 2009; Reinsel, Antrobus, & Wollman, 1992; Wamsley et al., 2007).
Based on this brief overview, we argue that dreaming is an involuntary form of embodied simulation that occurs whenever there is (1) an intact neural substrate that can support the process of dreaming, a qualification that allows for the impact of lesions on the functioning of this substrate; (2) an adequate level of cortical activation, which is provided by thalamic and extra-thalamic subcortical ascending pathways (e.g., Edlow et al., 2012; Jones, 2003; McGinty & Szymusiak, 2011) in waking and in some stages of sleep; (3) an occlusion of external stimuli; (4) a fully mature cognitive imagination system (as shown necessary by the apparent lack of dreaming in preschoolers and its relative paucity until ages 8--9); and (5) the loss of conscious self-control (Foulkes, 1999, pp. 124--125; Foulkes & Domhoff, 2014, p. 170).
Based on these specifications of what dreaming is and when it occurs, the remainder of this article suggests a way in which the neural network for dreaming might be examined in more detail by using waking mind-wandering, along with knowledge of its relation to the default network, as the starting point for a search for episodes of dreaming during periods of mind-wandering. To repeat for clarity's sake, dreaming is revealed by mentation reports that include the dreamer and other characters observing or participating in imagined activities and social interactions within settings that are usually specified, that is, a simulated social world. The involuntary form of embodied simulation called dreaming occurs when the five criteria outlined in the previous paragraph are met, which may happen during waking or sleep.
Dreaming while awake
Brief episodes of dreaming during periods of drifting waking thought were inadvertently discovered by Foulkes (1985; pp. 71--72) and his colleagues during practice sessions meant to prepare participants to focus on their most recent thoughts during sleep awakenings. In a laboratory study in which participants reclined in a moderately lighted room, with their wakefulness monitored by EEG and EMG recordings, Foulkes and Scott (1973) learned that 24 percent of sampled thoughts reported by 16 female college students were described as visual and dramatic, and experienced as dreams. In a replication study with 10 men and 10 women, who were signaled to report their thoughts at 12 random times in sessions of 45--60 min, Foulkes and Fleisher (1975) found a very similar figure for dreaming episodes: 19 percent. Another 20 percent contained "what might be called mind-wandering (the subject is not controlling his thoughts, but he is aware he is in the laboratory...)," and yet another 22 percent contained "what might be called 'lost in thought' (the subject may or may not be controlling his thoughts, but he is not aware he is in the laboratory. . .)" (Foulkes & Fleisher, 1975, p. 69). No instances of self-described or objective sleep were recorded during these studies (Foulkes & Fleisher, 1975, p. 70). These findings were replicated again as a preliminary part of a study of REM and NREM dream reports from 16 participants (Foulkes & Schmidt, 1983), but were not formally published (Foulkes, 1985, p. 89, footnote 39, and pp. 70--77 for a summary of all three studies). Since these studies found dreaming as well as the same types of drifting waking thought and mind-wandering that were later reported in studies of the default network, it seems plausible that similar probes in future studies might discover brief episodes of dreaming during mind-wandering as well (a possibility discussed in more detail in Section 7).
An enhanced default network: Dreaming as intensified mind-wandering
The main features of the default network have been known for over a decade (e.g., Buckner, 2012; Buckner, Andrews- Hanna, & Schacter, 2008; Greicius, Krasnow, Reiss, & Menon, 2003; Gusnard & Raichle, 2001; Raichle et al., 2001; Schacter et al., 2008). They include the medial prefrontal cortex (which in turn has several sub-regions), the medial temporal lobe, the bilateral inferior parietal lobule/temporoparietal junction, and the posterior cingulate cortex, with the medial prefrontal cortex (MFPC) and the posterior cingulate cortex (PCC) serving as major hubs. This network, which includes other areas discussed throughout this section, is one of three intrinsic networks within the overall network of extrinsic and intrinsic brain networks (Doucet et al., 2011, p. 2756; Fox et al., 2005; Sporns, 2011; Yeo et al., 2011; Yuan, Zotev, Phillips, Drevets, & Bodurka, 2012). In a human connectome study, it was found that "For all combinations of methods, areas of the default mode network showed the highest structure--function agreement within the whole brain. . ." (Horn, Ostwalda, Reisert, & Blankenburg, 2013).
Despite the apparent centrality of the default network in supporting drifting waking thought, a quantitative meta-analysis using activation likelihood estimation (ALE) (Eickhoff, Bzdok, Laird, Kurth, & Fox, 2012; Eickhoff et al., 2009) found that studies explicitly examining mind-wandering and related 'spontaneous' thought processes consistently recruited non-default network regions as well (Fox, Spreng, Ellamil, Andrews-Hanna, & Christoff, 2015). Using five stringent criteria for inclusion in the study, the meta-analysis of ten studies found 13 regions that were more active in spontaneous thought, as compared to various task-oriented comparison conditions with lower levels of mind-wandering. Prominent among the results were all the major hubs of the default network: the MPFC, the PCC, the medial temporal lobe, and bilateral inferior parietal lobule (Fox et al., 2015, Table 2 and Fig. 1). Other meta-analytic clusters included secondary somatosensory areas, which is consistent with the psychological literature on embodied cognition (Gibbs, 2006; Wilson, 2002). In addition, an area involved in visual imagery, the lingual gyrus, was also activated during spontaneous waking thought.
Research focused on the specific nature of the thinking supported by the default network has revealed two subsystems within it (Andrews-Hanna, Reidler, Sepulcre, Poulin, & Buckner, 2010). A dorsal medial subsystem is activated by instructions to think about the present situation or a present mental state ("present self"). It includes the dorsomedial prefrontal cortex, the temporoparietal junction/anterior inferior parietal lobule, the lateral temporal cortex, and temporopolar cortex (the anterior pole of the temporal lobe). The medial temporal subsystem is activated by thinking about personal situations and decisions in the future ("future self"); it includes the ventral medial prefrontal cortex, posterior inferior parietal lobule, retrosplenial cortex, parahippocampal cortex, and hippocampal formation (Andrews-Hanna, Reidler, Sepulcre, et al., 2010, p. 559). As found in earlier studies, "both subsystems are activated during passive states, when participants engage in spontaneous cognition" (Andrews-Hanna, Reidler, Sepulcre, et al., 2010, p. 559).
An activation likelihood estimation (ALE) meta-analysis (Eickhoff et al., 2009, 2012), based on 18 neuroimaging studies of the default network and six PET neuroimaging studies of the brain during REM sleep, showed that the two states showed differential patterns of brain activation, suggesting that "dreaming amplifies the same features that distinguish mind-wandering from goal-directed thought" (Fox et al., 2013, p. 1). In particular, the medial prefrontal cortex, the medial temporal lobe bilaterally (specifically, parahippocampal and entorhinal cortex), the lingual gyrus (also bilaterally), and the caudate nucleus were found to be more active in REM sleep, as compared to restful waking. Perhaps just as important, prefrontal areas that support executive functions, such as the orbitofrontal cortex and rostrolateral prefrontal cortex, were less active (Fox et al., 2015, Table 2). While first-person reports were only occasionally collected in the studies that were meta-analyzed, it is safe to assume that the REM periods were very likely accompanied by dreaming, based on previous laboratory findings that 80--90 percent of REM awakenings lead to dream reports (e.g., Dement, 1965; Dement & Kleitman, 1957). Similarly, every neuroimaging study to date of mind-wandering has shown strong involvement of the default network (e.g., Addis, Pan, Vu, Laiser, & Schacter, 2009; Andrews-Hanna, Reidler, Huang, et al., 2010; Christoff et al., 2009; Mason et al., 2007), so default network activations seem a fair proxy indicator for spontaneous thought. (Note, however, that our recent meta-analytic work has shown that the neural basis of mind-wandering involves more than just default network activity (Fox et al., 2015).)
Moreover, these findings are consistent with the results of lesion studies of dreaming. Neurological injuries to the areas that are inactive during REM, such as the lateral prefrontal cortex, the primary visual cortex, and primary sensorimotor cortices, do not affect dreaming according to patient reports (Solms, 1997). On the other hand, lesions in secondary visual cortex, which supports the generation of visual imagery, lead to the loss of visual imagery in both dreaming and waking, as documented in both sleep laboratory studies and clinical accounts (Kerr & Foulkes, 1981; Kerr, Foulkes, & Jurkovic, 1978; Solms, 1997, Chapters 11--13). Similarly, lesions in the grey or white matter of the ventral portion of MPFC, or in the area of the temporoparietal junction, lead to a global loss of dreaming (Bischof & Basset, 2004; Poza & Masso, 2006; Solms, 1997, Chapters 4 and 16; Yu, 2006).
Figure 1. Brain areas critical to both daydreaming and dreaming.
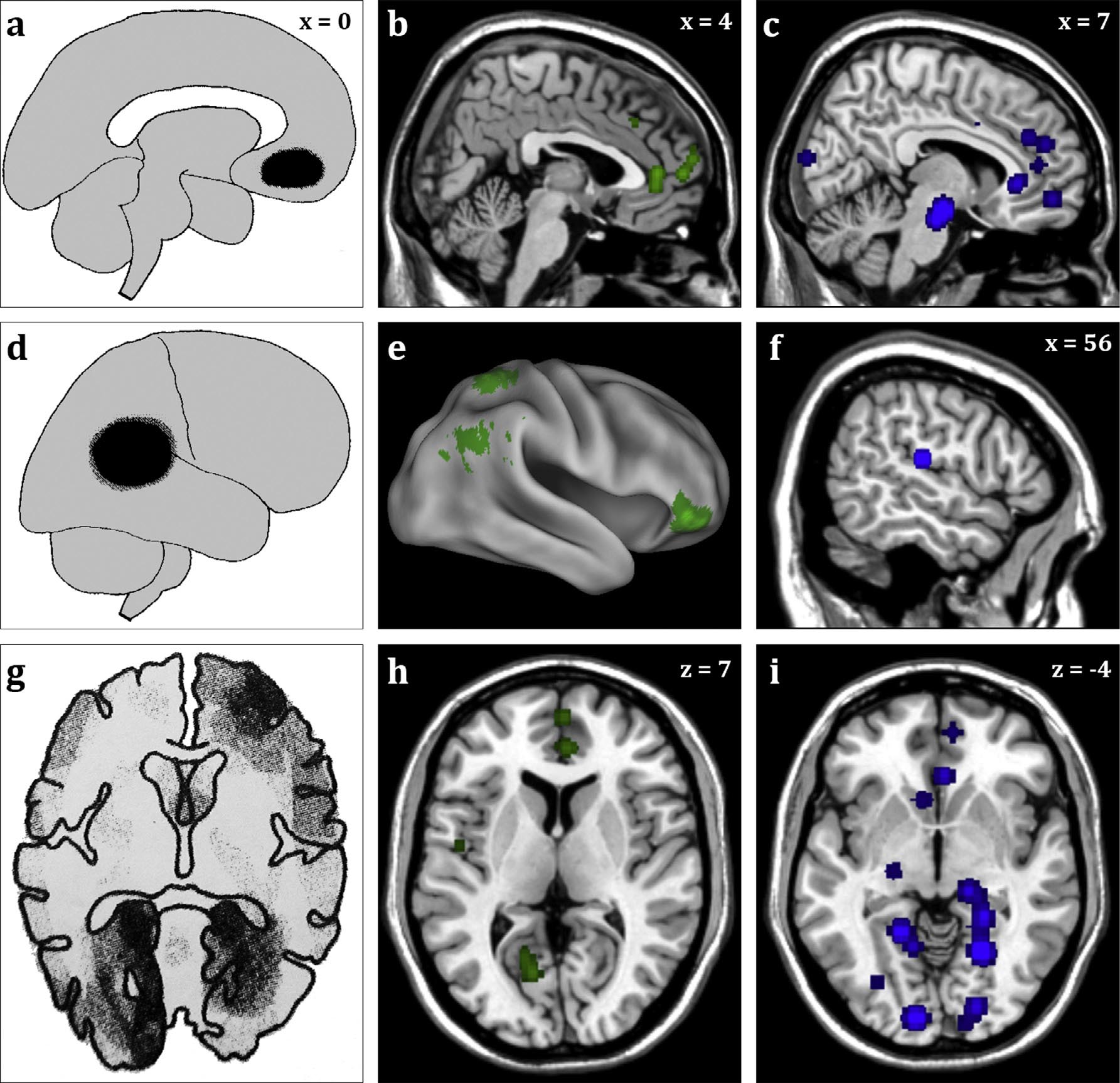 Note: The left column shows areas critical to dreaming as determined by overlapping lesion sites based on CT scans of neurological patients. The middle column shows meta-analytic brain activations associated with waking daydreaming/mindwandering. The right column shows meta-analytic brain activations associated with REM sleep, which is nearly always accompanied by dreaming. All three approaches converge on three areas as being central to dreaming: the medial prefrontal cortex (panels a-c); the temporoparietal junction/anterior inferior parietal lobule (panels d-f); and the medial occipital lobes/lingual gyrus for the visual aspects of dreaming in particular (panels g--i). Panels a and d modified, with permission, from Solms (2000); panel g modified with permission from Solms (1997). Panels b, e, and h based on data from Fox et al. (2015). Panels c, f, and i based on data from Fox et al. (2013). X and Z values represent left--right and vertical coordinates, respectively, in stereotactic space.
The overlap between the findings from neuropsychological lesion studies, and meta-analyses of neuroimaging studies during dreaming and daydreaming in healthy adults, is striking enough to warrant more detailed consideration (Figure 1). In his study of the dream mentation of hundreds of neurological patients with an enormous variety of lesions, Solms (1997) concluded that three broad brain areas were critical for dreaming: severe damage to either the ventral medial prefrontal cortex (Figure 1a) or the temporoparietal junction/anterior inferior parietal lobule (Figure 1d) led to a global cessation of dreaming, whereas damage to the secondary visual areas of the medial occipital lobe/lingual gyrus (Figure 1g) led to a cessation of visual dream imagery (while leaving dreaming otherwise normal). Our meta-analysis of brain activations during waking mind-wandering/daydreaming (Fox et al., 2015) shows significantly activated clusters in all three of these regions (Figure 1b, e, and h, respectively). Our meta-analysis of brain activity during REM sleep (as compared to quiet waking) similarly shows elevated activation in MPFC (Figure 1c), temporoparietal junction (Figure 1f), and medial occipital areas (Figure 1i). Our original meta-analysis of REM sleep activity (Fox et al., 2013), upon which the latter data are based, did not show significantly elevated activity in the temporoparietal junction/anterior inferior parietal lobule, but this was due to the use of stringent statistical thresholds in conjunction with the small sample size of the meta-analysis. Using slightly more lenient statistical thresholds, here, activations in this area are evident (Figure 1f).
A caveat to be kept in mind is that all three sets of data are based on comparatively small samples, resulting in reduced statistical power. Given how nascent this field of research is, the REM sleep meta-analysis was based on only 6 studies (Fox et al., 2013), and the mind-wandering meta-analysis on only 10 studies (Fox et al., 2015). Solms's (1997) neuropsychological lesion work was similarly based on very small groups of patients: overlap maps for the MPFC (Figure 1a) were based on 9 patients, for the temporoparietal junction (Figure 1d) on 8 patients, and for the medial occipital lobes (Figure 1g) on 6 patients (Solms, 1997). (Note, however, that despite the small number of cases used to actually create localization maps on brain templates, a large neuropsychological lesion literature further supports these findings (Solms, 2000).) Given these severe limitations in sample size, and the corresponding noise that must characterize these data sets, we find the overlap (Figure 1) all the more striking.
Although this overlap in the neural substrates of mind-wandering and dreaming is intriguing, it is precisely at this point that the type of study suggested in this article could come into play by making it possible to sort out the details of the similarities and differences in brain activation. Without going into further detail regarding these similarities and differences, there are two general findings that need to be highlighted in terms of the research agenda proposed in this article. First, the greater activation of regions such as the lingual gyrus (in the medial occipital lobe) and the caudate nucleus may help explain the enhanced sensorimotor qualities that are present in dreaming as compared to mind-wandering. For example, the lingual gyrus, a secondary visual area, appears to support the generation of visual imagery, and the caudate nucleus, part of the basal ganglia, is supportive of movement systems. These findings lead to the more specific hypothesis that dreaming is an intensified form of mind-wandering (Fox et al., 2013, p. 10) as well as being a form of embodied simulation. More precisely, intensified mind-wandering, in the context of low levels of activation in the regions that support executive, attentional, and primary sensorimotor networks, may make it possible for embodied simulations to be elevated into the dramatic enactments we call dreams (Fox et al., 2013; Klinger, 1990, 2009, for reviews of the differences between the content of daydreams and dreams).
Second, the greater activation of the MPFC during REM (almost certainly accompanied by dreaming) than during waking rest (very likely accompanied by mind-wandering) takes on added significance because there is evidence that this region may be the main source of the information flow among the main regions of the default network. One recent study concluded that "our structural results suggest that default network activity may be driven from. . . highly connected and central regions, such as the medial orbitofrontal cortex" (Hagmann et al., 2008, p. 0009). Similarly, Di and Biswal (2014, pp. 55--56, 58, Fig. 6) used dynamic causal modeling in a way that seems to obviate criticism of this method (Lohmann, Erfurth, Müller, & Turner, 2012), and thereby provided further evidence that the information flow in the default network largely originates in the MPFC. Also noteworthy in this context is that, whereas large lesions to ventral MPFC abolish dreaming entirely (see above), partial (often unilateral) lesions of the MPFC/anterior cingulate cortices can lead to an increased frequency and vividness of dreaming, or the intrusion of dreaming into waking thought, and a consequent confusion between dreaming and waking thought (Solms, 1997, Chapters 20--21; Whitty & Lewin, 1957). Another research group similarly found that their patients with lesions in these regions were suffering from "waking dreams" (Damasio, Graff-Radford, Eslinger, Damasio, & Kassell, 1985, p. 269).
Of perhaps even greater importance, the MPFC seems to be a central hub in the widely distributed waking self-system (e.g., Heatherton, 2010; Jenkins & Mitchell, 2011; Macrae, Moran, Heatherton, Banfield, & Kelley, 2004). Similarly, the default network shares many regions in common with the network for social cognition, which the authors of one review and synthesis call "an extended social-affective default network" (Amft et al., 2014, p. 1), and a meta-analysis has shown that the mentalizing network overlaps considerably with the default network as well (Spreng, Mar, & Kim, 2009). These results are consistent with the discovery that during mind-wandering "people tend to engage in self-relevant internal cognition processes predominantly about significant past and future events" (Andrews-Hanna, Reidler, Sepulcre, et al., 2010, p. 559); they also fit with the frequent focus on personal concerns and interpersonal interactions during dreaming (e.g., Domhoff, 1996, 2003, Chapter 5; Foulkes, 1985; Hall, 1951, 1953). Thus, if the MPFC in fact becomes an important driver during REM, then this may help to explain why dreaming as intensified mind-wandering is often focused on the dreamer's concerns about his or her relationships with significant others, regrets about the past, and worries about anxiety-arousing future events (Zadra & Domhoff, 2011).
For purposes of this article, the greater activation of areas such as the MPFC, the medial temporal lobe, the temporoparietal junction, the lingual gyrus, and the caudate nucleus during REM dreaming provide clues as to what areas might be more active during brief episodes of dreaming during extended periods of mind-wandering. However, there remains the need for a physiological indicator that could signal when brief episodes of dreaming are occurring during periods of mind-wandering that are being imaged with fMRI. Perhaps this indicator can be provided by changes in the EEG when secondary sensorimotor and/or subcortical areas suddenly augment the default network.
Electrophysiological indicators of default network activation
Several early EEG studies demonstrated that slow anterior alpha (in the 7--9 Hz range, with eyes closed) is associated with relaxed wakefulness (Cantero, Atienza, & Salas, 2002). Later EEG studies suggest that alpha is linked to the default network and spontaneous self-referential thinking (Knyazev, 2013; Knyazev, Slobodskoj-Plusnin, Bocharov, & Pylkova, 2011). Similarly, EEG/fMRI studies show that alpha is present when the default network is active, although alpha is not exclusive to that network (Jann et al., 2009; Mantini, Perrucci, Del Gratta, Romani, & Corbetta, 2007). Most significantly for our purposes, Chang, Liu, Chen, Liu, and Duyn (2013, p. 230) correlated EEG power with the degree of functional connectivity between and within resting-state networks, using participants who were instructed to keep their eyes closed. They discovered that alpha power is highest when the anterior default network (which importantly includes the ventromedial and anterior medial PFC) is most active while, simultaneously, key regions of the dorsal attention network are deactivated. This anti-correlated relationship between the default network and the dorsal attentional network has also been confirmed using a different regression methodology (Carbonell, Bellec, & Shmuel, 2014). In addition, Chang et al. (2013, p. 230) reported that "increased theta power was associated with increased functional connectivity between nodes of the [default network], particularly within anterior regions."
These various (and variously replicated) findings are important in any attempt to further specify the putative neural network for dreaming because they create bridges between fMRI studies of default network activation and the early EEG studies of dreaming during mind-wandering (discussed in Section 3). Moreover, the disconnection between the default network and the dorsal attention network is critical because decreased vigilance and the occlusion of external stimuli seem to be factors in setting the stage for dreaming (Antrobus et al., 1995; Foulkes, 1999; Foulkes & Domhoff, 2014). Then, too, the association of the default network and theta is noteworthy because both theta and alpha are present during sleep onset and in REM sleep (Badia, Wright, &Wauquier, 1994, pp. 215--216; Cantero et al., 2002; Hori, Hayashi, & Morikawa, 1994, p. 245; Marzano et al., 2011; Ogilvie, 2001).
Dreaming at sleep onset
The argument in this article, and the emphasis in the previous section on alpha as a possible indicator of default-network activation, gains added support from several different types of studies of dreaming during the sleep-onset process. This literature is of special interest because dreaming sometimes occurs before the EEG shows any signs of sleep, which once again demonstrates that dreaming can occur during a non-sleep state. Dreaming during sleep onset was demonstrated most clearly in the first systematic laboratory study of the sleep-onset process, which involved nine participants, eight men and one woman (Foulkes & Vogel, 1965). It was replicated with 32 participants, 16 women and 16 men (Foulkes, Spear, & Symonds, 1966). Based on answers to a series of questions asked immediately after the participants reported "what they were experiencing" when they were aroused, the researchers found an increasing number of dream reports in the first three stages in the four-stage process they identified, followed by a slight decline with the appearance of sleep spindles, which are the traditional indicators that a person is physiologically asleep. In addition, the increasing number of dream reports was accompanied by increasing drowsiness from stage to stage and a concomitant decline in the participants' belief that they were controlling their thoughts.
Moreover, even though there are more "fleeting progressions of visual imagery or dissociated images and thoughts" during sleep onset than during REM periods, the "typical" report during the sleep-onset period was a dream that was "no less well organized than its REM-period counterpart" (Foulkes & Vogel, 1965, p. 238). That is, once participants no longer subjectively felt they were awake and in control of their thoughts, most of their reports seemed to be based on the process of dreaming even though they are not yet formally asleep based on traditional criteria: "Dreaming thus occurs in the absence of REMs, emergent Stage 1, or any kind of Stage 1 altogether. . ." (Foulkes & Vogel, 1965, p. 239). The researchers conclude that the frequency of hypnagogic phenomena during the sleep-onset process is overestimated in comparison to the frequency of dreaming due to unrepresentative sampling (Foulkes & Vogel, 1965, p. 242).
Although nearly every participant went through the same general process (increased drowsiness, a declining sense of volitional control, and an increase in dreaming reports), there were wide individual differences in the pace at which the participants moved through these subjective changes. For example, five participants dreamt during the first two stages, both of which include alpha patterns, but four participants only dreamt during the last two stages, which have theta waves similar to those during REM sleep. Importantly, six of the nine participants had one or more dreams during periods of "more or less continuous alpha" (Foulkes & Vogel, 1965, p. 237). The appearance of dreaming during a descending alpha state dovetails with the finding of dreaming during alpha in the waking state, as discussed in Section 3. In addition, participants only reported dreams if they also reported a loss of volitional control, whatever the sleep-onset stage, which is noteworthy because loss of volitional control appears to be another condition necessary for dreaming to occur (Foulkes & Domhoff, 2014, p. 170).
The rapidity with which the transition to dreaming can occur during sleep onset, whether in daytime studies in the laboratory or at participants' usual bedtime, is demonstrated in a sleep-lab pilot study in which participants were able to continue to talk as they fell asleep. Verbalization under these conditions was practicable because auditory feedback from their voices was occluded by the use of white noise and visual input was blocked by eye coverings (Bertini, Lewis, & Witkin, 1964, p. 495). The recording of participants' ramblings made it possible in a few cases to hear them mention the appearance of visual imagery (Bertini et al., 1964, pp. 505--506).
A series of extremely detailed EEG studies of the sleep-onset process replicated and extended previous work (Hayashi, Katoh, & Hori, 1999; Hori, Hayash, & Hibino, 1992; Hori, Hayash, & Morikawa, 1990; Morikawa, Hayash, & Hori, 2002). The investigation team began by examining 5-s epochs in the EEG records of 26 participants ages 21--26, 14 males and 12 females, which were collected on two nonconsecutive nights. This analysis and a subsequent replication eventually resulted in an eight-stage process defined by a topographic EEG map (Tamaki, Nittono, Hayashi, & Hori, 2005). The first three stages are characterized by varying percentages of alpha, the next four by the presence of theta and a vertex sharp wave, and the final stage by the presence of sleep spindles (Hori et al., 1994, p. 245).
Each successive stage showed a decline in reaction time and in the subjective experience of feeling awake. Conversely, the anterior alpha/posterior alpha ratio (the A/P ratio) increased at each stage of the sleep-onset process. This anteriorization of alpha during sleep onset (now sometimes called "frontalization") was also discovered in three other EEG studies without awakenings (DeGennaro, Ferrara, Curcio, & Cristiani, 2001; DeGennaro et al., 2004; Hasan & Broughton, 1994). The declining frequency and anteriorization of alpha during sleep onset may be comparable to the presence of slow anterior alpha during relaxed wakefulness (Cantero et al., 2002).
Hori et al. (1994, p. 250) concluded that the subjective reports they obtained from their participants "fundamentally confirmed" the findings from the first laboratory study of dreaming at sleep onset (Foulkes & Vogel, 1965). However, they label the reports they obtained as "hypnagogic imagery," defined as the reporting of any visual, sensory, or kinesthetic imagery. Thus, it cannot be certain that their participants were always reporting "dream" experiences as defined here, even though one or more of these forms of imagery usually occur in a dream. With that caveat in mind, they found that reports of these hypnagogic experiences gradually increased from 23.3 percent, 25.8 percent, and 33.5 percent in the three alpha stages to a high of 45.7 percent during a stage defined by theta (Hori et al., 1990, p. 247, Table 3). This finding parallels their results concerning the decline in the perceived awake percent and the rise in the A/P ratio; however, the percentage of "hypnogogic" reports then gradually declined to 31.9 percent in the stage defined by sleep spindles (Hori et al., 1994, p. 247, Table 3). A replication study (Hayashi et al., 1999, p. 677, Table 1) on subjective reports from various sleep-onset stages produced generally similar results, 29.8 percent for stages characterized by alpha, 36.2 percent for stages characterized by theta, and 22.8 percent for the sleep-spindles stage.
Turning to other sources to add to this emerging general picture, EEG/fMRI studies show that the default network remains somewhat active during the sleep onset process and into NREM 2 (e.g., Horovitz, 2008; Larson-Prior et al., 2009; Laufs et al., 2007). However, and importantly, the anterior and posterior portions of the default network show decreased connectivity with one another, and the anterior portion of the default network decouples from the dorsal attentional network as well (Sämann et al., 2011; Tagliazucchi et al., 2013). These findings are similar to the EEG results reported by Foulkes and his co-workers (Foulkes & Vogel, 1965; Foulkes et al., 1966; Vogel, Foulkes, & Trosman, 1966) and by Hori and his co-workers (Hayashi et al., 1999; Hori et al., 1994; Morikawa et al., 2002; Tamaki et al., 2005).
Taken together, these several different types of studies of the sleep-onset process (some with awakenings, some based on EEG recordings without awakenings, and some using EEG/fMRI without awakenings) tentatively suggest it may be the default network that is supporting the episodes of dreaming that occur at varying points during the complex sleep-onset process. As already noted, this conclusion provides further support for the possibility that there can be dreaming during non-sleep states. And if the MPFC and other regions of the default network are at the heart of the neural network for dreaming, as suggested by work discussed in Section 3 (Andrews-Hanna, Reidler, Sepulcre, et al., 2010; Di & Biswal, 2014; Fox et al., 2013; Hagmann et al., 2008), then its decoupling from the dorsal attentional network may provide the neural basis for the loss of conscious self-control that is the final necessary condition for dreaming, as discussed at the end of Section 2 (Foulkes & Domhoff, 2014, p. 170).
A counterintuitive research agenda
If the neural network that supports dreaming is centered in the dorsal medial subsystem of the default network outlined by Andrews-Hanna, Reidler, Sepulcre, et al. (2010), and if it becomes ascendant whenever there is a decline in vigilance and a loss of volitional control, then there is reason to believe that people can indeed drift into dreaming during periods of relaxed wakefulness and mind-wandering. Based on the review and synthesis of the many converging avenues of empirical work we have discussed above, an attempt to further specify the neural network that supports dreaming might include the following steps.
Both a 'predictive' and/or a 'post hoc' approach could be taken. A 'post hoc' paradigm could monitor brain activity continuously with EEG/fMRI, administering thought probes quasi-randomly to determine the intensity and quality of the subject's thinking. Post-hoc sorting of EEG/fMRI data, based on the 'immersiveness,' intensity, or 'dreaminess' of participants' first-person reports, could determine whether elevated neural markers of the default network indeed correlate with instances of dreaming while awake, as our hypothesis predicts.
A study investigating a slightly different research question has found tentative support for this point. A recent study by Vanhaudenhuyse et al. (2011) investigated the involvement of three key default network regions in the 'intensity' of internal (vs. external, i.e. perceptual) awareness. They defined internal awareness as "all environmental stimuli-independent thoughts (e.g., inner speech, autobiographical memories, or wandering thoughts)." During continuous fMRI acquisition, subjects were prompted by quasi-random experience-sampling probes that asked them to rate the contents of their awareness on a 4-point scale from strongly external ('1') or moderately external ('2') through to moderately internal ('3') and strongly internal ('4'). Investigating three key default network regions-of-interest (the medial prefrontal cortex, posterior cingulate cortex, and bilateral medial temporal lobe), they found that activity in these regions correlated positively with the self-reported intensity of internal awareness (Vanhaudenhuyse et al., 2011). This study did not investigate the rest of the brain, and so cannot speak to the involvement of other areas of our proposed augmented default network for dreaming. Moreover, their definition of 'strong internal awareness' is not exactly what we mean by dreaming during waking -- although there are of course similarities, with dreaming presumably representing the extreme pole of the continuum of 'internal awareness' during waking. So although the study by Vanhaudenhuyse et al. (2011) is not a direct investigation or confirmation of our hypothesis, we consider it encouraging preliminary evidence in its favor.
In addition to such post hoc analyses, a complementary 'predictive' method could be employed as an even stronger test of our hypothesis. The process could begin with uninterrupted EEG/fMRI mappings of the default network in several participants who are asked to let their minds wander, with eyes closed, for 10--15 min. Once individual baselines and variations in alpha and theta frequencies were established, and perhaps making use of any changes in the A/P ratio, participants could be asked during periods of low-frequency anterior alpha, and/or mixed periods of low-frequency alpha and theta, what they were experiencing at the moment their name is called. fMRI data could also be monitored in real-time, and elevated activations in one or more default network regions used as a criterion for initiating thought-sampling probes. Our hypothesis predicts that higher default network activity, as indicated either by EEG or fMRI measures, or possibly both in combination, would predict more immersive and dream-like mental activity during waking. If participants report dream narratives and imagery, the fMRI results then could be analyzed in greater detail to determine which parts of the default network most strongly predicted reports of dreaming during waking, along with the degree to which secondary visual areas (e.g., lingual gyrus) and subcortical motor regions (e.g., caudate nucleus) were active. It might even be that at least some participants could be asked to think out loud while visual and auditory input and feedback were blocked, which might make it easier to discover any sudden augmentation of the default network during the transition from thought-like mind-wandering to the onset of dreaming. In addition, visual and auditory occlusions might render the fMRI scanner's background noise less of a distraction.
As mentioned in the introduction to this article, such studies might be carried out as an additional component to studies of the default network that were being conducted for a variety of primary purposes, perhaps during the process of acclimating participants at the outset of the main study. Such add-on studies in several neuroimaging laboratories might make the findings more robust, especially given the wide range of individual differences in generating and reporting fleeting subjective experiences.
Acknowledgments
Our thanks to Daniel Bissell for research assistance, David Foulkes for many helpful substantive and editorial suggestions, Kalina Christoff for many inspiring discussions, and two anonymous reviewers for asking us to expand, elaborate, or clarify our argument on several important issues.
References
Addis, D., Pan, L., Vu, M.-A., Laiser, N., & Schacter, D. (2009). Constructive episodic simulation of the future and the past: Distinct subsystems of a core brain network mediate imagining and remembering. Neuropsychologia, 47, 2222-2238.
Addis, D., Wong, A. T., & Schacter, D. (2007). Remembering the past and imagining the future: Common and distinct neural substrates during event construction and elaboration. Neuropsychologia, 45, 1363-1377.
Amft, M., Bzdok, D., Laird, A., Fox, P., & Schilbach, L. (2014). Definition and characterization of an extended social-affective default network. Brain Structure & Function, PMID: 24399179 (PMCID: PMC4087104), [Available on 2015/2017/2018].
Andrews-Hanna, J. R. (2012). The brain's default network and its adaptive role in internal mentation. The Neuroscientist, 18, 251-270.
Andrews-Hanna, J. R., Reidler, J., Huang, C., & Buckner, R. (2010). Evidence for the default network's role in spontaneous cognition. Journal of Neurophysiology, 104, 322-335.
Andrews-Hanna, J. R., Reidler, J., Sepulcre, J., Poulin, R., & Buckner, R. (2010). Functional-anatomic fractionation of the brain's default network. Neuron, 65, 550-562.
Andrews-Hanna, J. R., Smallwood, J., & Spreng, R. N. (2014). The default network and self-generated thought: Component processes, dynamic control, and clinical relevance. Annals of the New York Academy of Science. http://dx.doi.org/10.1111/nyas.12360 (PMID: 24502540), PMC4039623 [Available on 4032015/4039625/4039621].
Antrobus, J., Kondo, T., & Reinsel, R. (1995). Dreaming in the late morning: Summation of REM and diurnal cortical activation. Consciousness & Cognition, 4, 275-299.
Badia, P., Wright, K., & Wauquier, A. (1994). Fluctations in single-hertz EEG activity during the transition to sleep. In R. Ogilvie & J. Harsh (Eds.), Sleep onset: Normal and abnormal processes (pp. 201-218). Washington: American Psychological Association.
Bergen, B. (2012). Louder than words: The new science of how the mind makes meaning. New York: Basic Books.
Bertini, M., Lewis, H., & Witkin, H. (1964). Some preliminary observations with an experimental procedure for the study of hypnagogic and related phenomena. Archivo Di Psicologia, Neurologia e Psichiatria, 6, 493-534.
Bischof, M., & Basset, C. (2004). Total dream loss: A distinct neuropsychological dysfunction after bilateral PCA stroke. Annals of Neurology, 56, 583-586.
Braun, A., Balkin, T., Wesensten, N., Carson, R., Varga, M., Baldwin, P., et al (1997). Regional cerebral blood flow throughout the sleep-wake cycle: An (H2O)- O-15 PET study. Brain, 120, 1173-1197.
Buckner, R. (2012). The serendipitous discovery of the brain's default network. NeuroImage, 62, 1137-1145.
Buckner, R., Andrews-Hanna, J. R., & Schacter, D. (2008). The brain's default network: Anatomy, function, and relevance to disease. Annals of the New York Academy of Sciences, 1124, 1-38.
Cantero, J., Atienza, M., & Salas, R. (2002). Human alpha oscillations in wakefulness, drowsiness period, and REM sleep: Different electroencephalographic phenomena within the alpha band. Neurophysiological Clinical, 32, 54-71.
Carbonell, F., Bellec, P., & Shmuel, A. (2014). Quantification of the impact of a confounding variable on functional connectivity confirms anti-correlated networks in the resting-state. NeuroImage, 86, 343-353.
Chang, C., Liu, Z., Chen, M., Liu, X., & Duyn, J. (2013). EEG correlates of time-varying BOLD functional connectivity. NeuroImage, 72, 227-236.
Chow, H. M., Horovitz, S., Carr, W., Picchioni, D., Coddington, N., Fukunaga, M., et al (2013). Rhythmic alternating patterns of brain activity distinguish rapid eye movement sleep from other states of consciousness. Proceedings of the National Academy of Sciences, 110, 10300-10305.
Christoff, K., Gordon, A., Smallwood, J., Smith, R., & Schooler, J. (2009). Experience sampling during fMRI reveals default network and executive system contributions to mind wandering. Proceedings of the National Academy of Sciences of the United States of America, 106, 8719-8724.
Cicogna, P., Natale, V., Occhionero, M., & Bosinelli, M. (1998). A comparison of mental activity during sleep onset and morning awakening. Sleep, 21(5), 462-470.
Damasio, A., Graff-Radford, N., Eslinger, P., Damasio, H., & Kassell, N. (1985). Amnesia following basal forebrain lesions. Archives of Neurology, 42, 263-271.
De Havas, J., Parimal, S., Soon, C., & Chee, M. (2012). Sleep deprivation reduces default mode network connectivity and anti-correlation during rest and task performance. NeuroImage, 59, 1745-1751.
DeGennaro, L., Ferrara, M., Curcio, G., & Cristiani, R. (2001). Antero-posterior EEG changes during the wakefulness-sleep transition. Clinical Neurophysiology, 112, 1901-1911.
DeGennaro, L., Vecchio, F., Ferrara, M., Curcio, G., Rossini, P., & Babiloni, C. (2004). Changes in fronto-posterior functional coupling at sleep onset in humans. Journal of Sleep Research, 13, 209-217.
Dement, W. (1965). An essay on dreams: The role of physiology in understanding their nature. In T. Newcomb (Ed.), New directions in psychology II (pp. 135-257). New York: Holt, Rinehart, and Winston.
Dement, W., & Kleitman, N. (1957). The relation of eye movements during sleep to dream activity: An objective method for the study of dreaming. Journal of Experimental Psychology, 53, 339-346.
Di, X., & Biswal, B. (2014). Identifying the default mode network structure using dynamic causal modeling on resting-state functional magnetic resonance imaging. NeuroImage, 86, 53-59.
Dixon, M. L., Fox, K. C. R., & Christoff, K. (2014). A framework for understanding the relationship between externally and internally directed cognition. Neuropsychologia, 62, 321-330.
Domhoff, G. W. (1996). Finding meaning in dreams: A quantitative approach. New York: Plenum.
Domhoff, G. W. (2001). A new neurocognitive theory of dreams. Dreaming, 11, 13-33.
Domhoff, G. W. (2003). The scientific study of dreams: Neural networks, cognitive development, and content analysis. Washington, DC: American Psychological Association.
Domhoff, G. W. (2011a). Dreams are embodied simulations that dramatize conceptions and concerns: The continuity hypothesis in empirical, theoretical, and historical context. International Journal of Dream Research, 4, 50-62.
Domhoff, G. W. (2011b). The neural substrate for dreaming: Is it a subsystem of the default network? Consciousness and Cognition, 20, 1163-1174.
Domhoff, G. W., & Schneider, A. (1999). Much ado about very little: The small effect sizes when home and laboratory collected dreams are compared. Dreaming, 9, 139-151.
Doucet, G., Naveau, M., Petit, L., Delcroix, N., Zago, L., Crivello, F., et al (2011). Brain activity at rest: A multiscale hierarchical functional organization. Journal of Neurophysiology, 105, 2753-2763.
Edlow, B., Takahashi, E., Wu, O., Benner, T., Dai, G., Bu, L., et al (2012). Neuroanatomic connectivity of the human ascending arousal system critical to consciousness and its disorders. Journal of Neuropathology and Experimental Neurology, 71, 531-546.
Eickhoff, S., Bzdok, D., Laird, A., Kurth, F., & Fox, P. (2012). Activation likelihood estimation meta-analysis revisited. NeuroImage, 59, 2349-2361.
Eickhoff, S., Laird, A., Grefkes, C., Wang, L., Zilles, K., & Fox, P. (2009). Coordinate-based activation likelihood estimation meta-analysis of neuroimaging data: A random-effects approach based on empirical estimates of spatial uncertainty. Human Brain Mapping, 30, 2907-2926.
Foulkes, D. (1985). Dreaming: A cognitive-psychological analysis. Hillsdale, NJ: Erlbaum.
Foulkes, D. (1999). Children's dreaming and the development of consciousness. Cambridge, MA: Harvard University Press.
Foulkes, D., & Domhoff, G. W. (2014). Bottom-up or top-down in dream neuroscience? A top-down critique of two bottom-up studies. Consciousness and Cognition, 27, 168-171.
Foulkes, D., & Fleisher, S. (1975). Mental activity in relaxed wakefulness. Journal of Abnormal Psychology, 84, 66-75.
Foulkes, D., & Schmidt, M. (1983). Temporal sequence and unit comparison composition in dream reports from different stages of sleep. Sleep, 6, 265-280.
Foulkes, D., & Scott, E. (1973). An above-zero baseline for the incidence of momentarily hallucinatory mentation. Sleep Research, 2, 108.
Foulkes, D., Spear, P., & Symonds, J. (1966). Individual differences in mental activity at sleep onset. Journal of Abnormal and Social Psychology, 71, 280-286.
Foulkes, D., & Vogel, G. (1965). Mental activity at sleep onset. Journal of Abnormal Psychology, 70, 231-243.
Fox, K. C. R., Nijeboer, S., Solomonova, E., Domhoff, G. W., & Christoff, K. (2013). Dreaming as mind wandering: Evidence from functional neuroimaging and first-person content reports. Frontiers in Human Neuroscience, 7(Article 412), 118. http://dx.doi.org/10.3389/fnhum.2013.00412 (eCollection 02013).
Fox, K. C. R., Spreng, R. N., Ellamil, M., Andrews-Hanna, J. R., & Christoff, K. (2015). The wandering brain: Meta-analysis of functional neuroimaging studies of mind-wandering and related spontaneous thought processes. NeuroImage (in press).
Fox, M., Snyder, A., Vincent, J., Corbetta, M., Van Essen, D., & Raichle, M. (2005). The human brain is intrinsically organized into dynamic, anticorrelated functional networks. Proceedings of the National Academy of Sciences, 102, 9673-9678.
Gibbs, R. (2006). Embodiment and cognitive science. New York: Cambridge University Press.
Gibbs, R. (2014). Conceptual metaphor in thought and social action. In M. Landau, M. Robinson, & B. Meier (Eds.), The power of metaphor: Examining its influence on social life (pp. 17-40). Washington: American Psychological Association.
Greicius, M., Krasnow, B., Reiss, A., & Menon, V. (2003). Functional connectivity in the resting brain: A network analysis of the default mode hypothesis. Proceedings of the National Academy of Sciences, 100, 253-258.
Gusnard, D., & Raichle, M. (2001). Searching for the baseline: Functional imagining and the resting human brain. Nature Reviews Neuroscience, 2, 685-694.
Hagmann, P., Commoun, L., Gigandet, X., Meuli, R., Honey, C., Wedeen, V., et al (2008). Mapping the structural core of human cerebral cortex. PLoS Biology, 6(7). http://dx.doi.org/10.1371/journal.pbio.0060159.
Hall, C. (1951). What people dream about. Scientific American, 184, 60-63.
Hall, C. (1953). A cognitive theory of dreams. Journal of General Psychology, 49, 273-282.
Hall, C., & Van de Castle, R. (1966). The content analysis of dreams. New York: Appleton-Century-Crofts.
Hasan, J., & Broughton, R. (1994). Quantitative topographic EEG mapping during drowsiness and sleep. In R. Ogilvie & J. Harsh (Eds.), Sleep onset: Normal and abnormal processes (pp. 219-235). Washington: American Psychological Association.
Hayashi, M., Katoh, K., & Hori, T. (1999). Hypnagogic imagery and EEG activity. Perceptual and Motor Skills, 88, 676-678.
Heatherton, T. (2010). Building a social brain. In P. Reuter-Lorenz, K. Baynes, G. Mangun, & E. Phelps (Eds.), The cognitive neuroscience of mind (pp. 173-188). Cambridge: MIT Press.
Hobson, J. A. (2002). Dreaming: An introduction to the science of sleep. New York: Oxford University Press.
Hori, T., Hayash, i. M., & Morikawa, T. (1990). Typography and coherence analysis of hypnagogic EEG. In J. Horne (Ed.), Sleep '90 (pp. 10-12). Bochum, Germany: Pontenagel Press.
Hori, T., Hayash, M., & Hibino, K. (1992). An EEG study of the hypnagogic hallucinatory experience. International Journal of Psychology, 27, 420.
Hori, T., Hayashi, M., & Morikawa, T. (1994). Topographic EEG changes and the hypnagogic experience. In R. Ogilvie & J. Harsh (Eds.), Sleep onset: Normal and abnormal processes. Washington: American Psychological Association.
Horn, A., Ostwalda, D., Reisert, M., & Blankenburg, F. (2013). The structural-functional connectome and the default mode network of, the human brain. NeuroImage, 102, 142-151.
Horovitz, S. (2008). Low frequency BOLD fluctuations during resting wakefulness and light sleep: A simultaneous EEG-fMRI study. Human Brain Mapping, 29, 671-682.
Ioannides, A., Kostopoulos, K., Liu, L., & Fenwick, P. (2009). MEG identifies dorsal medial brain activations during sleep. NeuroImage, 44, 455-468.
Jann, K., Dierks, T., Boesch, C., Kottlow, M., Strik, W., & Koenig, T. (2009). BOLD correlates of EEG alpha phase-locking and the fMRI default mode network. NeuroImage, 45, 903-916.
Jenkins, A., & Mitchell, J. (2011). Medial prefrontal cortex subserves diverse forms of self-reflection. Social Neuroscience, 6, 211-218.
Jones, B. E. (2003). Arousal systems. Frontiers in Bioscience, 8, 438-451.
Kass, W., Preiser, G., & Jenkins, A. (1970). Inter-relationship of hallucinations and dreams in spontaneously hallucinating patients. Psychiatric Quarterly, 44(3), 488-499.
Kerr, N., & Foulkes, D. (1981). Right hemispheric mediation of dream visualization: A case study. Cortex, 17, 603-610.
Kerr, N., Foulkes, D., & Jurkovic, G. (1978). Reported absence of visual dream imagery in a normally sighted subject with Turner's syndrome. Journal of Mental Imagery, 2, 247-264.
Klinger, E. (1990). Daydreaming. Los Angeles: Jeremy P. Tarcher.
Klinger, E. (2009). Daydreaming and fantasizing: Thought flow and motivation. In K. Markman, W. Klein, & J. Suhr (Eds.), Handbook of imagination and mental simulation (pp. 225-239). New York: Psychology Press.
Knyazev, G. (2013). EEG correlates of self-referential processing. Frontiers in Human Neuroscience, 7, 264. http://dx.doi.org/10.3389/ fnhum.2013.00264eBook.
Knyazev, G., Slobodskoj-Plusnin, J., Bocharov, A., & Pylkova, L. (2011). The default mode network and EEG alpha oscillations: An independent component analysis. Brain Research, 1402, 67-79.
Kucyi, A., & Davis, K. D. (2014). Dynamic functional connectivity of the default mode network tracks daydreaming. NeuroImage, 100, 471-480.
Larson-Prior, L., Zempel, J., Nolan, T., Prior, F., Snyder, A., & Raichle, M. (2009). Cortical network functional connectivity in the descent to sleep. Proceedings of the National Academy of Sciences of the United States, 106, 4489-4494.
Laufs, H. (2008). Endogenous brain oscillations and related networks detected by surface EEG-combined fMRI. Human Brain Mapping, 29, 762-769.
Laufs, H., Walker, M., & Lund, T. (2007). Brain activation and hypothalamic functional connectivity during human non-rapid eye movement sleep: An EEG/ fMRI study-Its limitations and an alternative approach. Brain, 130, 75-84.
Lauro, L., Mattavelli, G., Papagno, C., & Tettamanti, M. (2013). She runs, the road runs, my mind runs, bad blood runs between us: Literal and figurative motion verbs, an fMRI study. NeuroImage, 83, 361-371.
Lohmann, G., Erfurth, K., Müller, K, & Turner, R. (2012). Critical comments on dynamic causal modelling. NeuroImage, 59, 2322-2329.
Macrae, C. N., Moran, J. M., Heatherton, T. F., Banfield, J. F., & Kelley, W. M. (2004). Medial prefrontal activity predicts memory for self. Cerebral Cortex, 14, 647-654.
Mantini, D., Perrucci, M., Del Gratta, C., Romani, G., & Corbetta, M. (2007). Electrophysiological signatures of resting state networks in the human brain. Proceedings of the National Academy of Sciences, 104, 13170-13175.
Maquet, P., Peters, J., Aerts, J., Delfiore, G., Dequerldre, C., Luxen, A., et al (1996). Functional neuroanatomy of human rapid-eye-movement sleep and dreaming. Nature, 383, 163-166.
Marzano, C., Ferrara, M., Mauro, F., Moroni, F., Gorgoni, M., Tempesta, D., et al (2011). Recalling and forgetting dreams: Theta and alpha oscillations during sleep predict subsequent dream recall. Journal of Neuroscience, 31, 6647-6683.
Mason, M., Norton, M., Van Horn, J., Wenger, D., Grafton, S., & Macrae, N. (2007). Wandering minds: The default network and stimulus-independent thought. Science, 315, 393-395.
McGinty, D., & Szymusiak, R. (2011). Neural control of sleep in mammals. In M. Kryger, T. Roth, &W. Dement (Eds.), Principles and practices of sleep medicine (5th ed., pp. 76-91). Philadephia: Elsevier Saunders.
Morikawa, T., Hayash, i. M., & Hori, T. (2002). Spatiotemporal variations of alpha and sigma band EEG in the waking-sleeping transition period. Perceptual and Motor Skills, 95, 131-154.
Niedenthal, P., Winkielman, P., Mondillon, L., & Vermeulen, N. (2009). Embodiment of emotion concepts. Journal of Personality and Social Psychology, 96, 1120-1136.
Nielsen, T. (2000). A review of mentation in REM and NREM sleep: "Covert" REM sleep as a possible reconciliation of two opposing models. Behavioral and Brain Sciences, 23, 851-866.
Nofzinger, E., Mintun, M., Wiseman, M., Kupfer, D., & Moore, R. (1997). Forebrain activation in REM sleep: An FDG PET study. Brain Research, 770, 192-201.
Nofzinger, E., Nichols, T., Meltzer, C., Price, J., Steppe, D., Miewald, J., et al (1999). Changes in forebrain function from waking to REM sleep in depression: Preliminary analyses of (18F)FDG PET studies. Psychiatry Research, 91(2), 59-78.
Ogilvie, R. (2001). The process of falling asleep. Sleep Medicine Review, 5, 247-270.
Pagel, J. F., Blagrove, M., Levin, R., States, B., Stickgold, B., & White, S. (2001). Definitions of dream: A paradigm for comparing field descriptive specific studies of dream. Dreaming, 11(4), 195-202.
Poza, I., & Masso, J. (2006). Total dream loss secondary to left temporo-occipital brain injury. Neurologia, 21, 152-154.
Raichle, M., MacLeod, A., Snyder, A., Powers, W., Gusnard, D., & Shulman, G. (2001). A default mode of brain function. Proceedings of the National Academy of Science, 98, 676-682.
Reinsel, R., Antrobus, J., & Wollman, M. (1992). Bizarreness in dreams and waking fantasy. In J. Antrobus & M. Bertini (Eds.), The neuropsychology of sleep and dreaming (pp. 157-184). Hillsdale, NJ: Erlbaum.
Sacks, O. (2013). Hallucinations. New York: Knopf.
Sämann, P., Wehrle, R., Hoehn, D., Spoormaker, V., Peters, H., Tully, C., et al (2011). Development of the brain's default mode network from wakefulness to slow wave sleep. Cerebral Cortex, 21, 2082-2093.
Schacter, D., Addis, D., & Buckner, R. (2008). Episodic simulation of future events: Concepts, data, and applications. Annals of the New York Academy of Sciences, 1124, 39-60.
Solms, M. (1997). The neuropsychology of dreams: A clinico-anatomical study. Hillsdale, NJ: Erlbaum.
Solms, M. (2000). Dreaming and REM sleep are controlled by different brain mechanisms. Behavioral and Brain Sciences, 23, 843-850.
Solms, M., & Turnbull, O. (2002). The brain and the inner world. New York: Other Press.
Sporns, O. (2011). Networks of the brain. Cambridge: MIT Press.
Spreng, R. N., Mar, R., & Kim, A. (2009). The common neural basis of autobiographical memory, prospection, navigation, theory of mind, and the default mode: A quantitative meta-analysis. Journal of Cognitive Neuroscience, 21, 489-510.
Tagliazucchi, E., von Wegner, F., Morzelewski, A., Brodbeck, V., Jahnke, K., & Laufs, H. (2013). Breakdown of long-range temporal dependence in default mode and attention networks during deep sleep. Proceedings of the National Academy of Science, 110, 15419-15424.
Tamaki, M., Nittono, H., Hayashi, M., & Hori, T. (2005). Examination of the first-night effect during the sleep-onset period. Sleep, 28, 195-202.
Vanhaudenhuyse, A., Demertzi, A., Schabus, M., Noirhomme, Q., Bredart, S., Boly, M., et al (2011). Two distinct neuronal networks mediate the awareness of environment and of self. Journal of cognitive neuroscience, 23, 570-578.
Vogel, G., Foulkes, D., & Trosman, H. (1966). Ego functions and dreaming at sleep onset. Archives of General Psychiatry, 14, 238-248. Wamsley, E., Hirota, Y., Tucker, M., Smith, M., Doan, T., & Antrobus, J. (2007). Circadian and ultradian influences on dreaming: A dual rhythm model. Brain Research Bulletin, 71, 347-354.
Whitty, C., & Lewin, W. (1957). Vivid day-dreaming: An unusual form of confusion following anterior cingulectomy. Brain, 80, 72-76.
Wilson, M. (2002). Six views of embodied cognition. Psychonomic Bulletin & Review, 9, 625-636.
Yeo, B., Krienen, F., Sepulcre, J., Sabuncu, M., Lashkari, D., Hollinshead, M., et al (2011). The organization of the human cerebral cortex estimated by intrinsic functional connectivity. Journal of Neurophysiology, 106, 1125-1165.
Yu, C. (2006). The brain mechanims of dreaming. Unpublished doctoral dissertation, University of Cape Town, Cape Town, South Africa.
Yuan, H., Zotev, V., Phillips, R., Drevets, W., & Bodurka, J. (2012). Spatiotemporal dynamics of the brain at rest: Exploring EEG microstates as electrophysiological signatures of BOLD resting state networks. NeuroImage, 60, 2062-2072.
Zadra, A., & Domhoff, G. W. (2011). The content of dreams: Methods and findings. In M. Kryger, T. Roth, & W. Dement (Eds.), Principles and practices of sleep medicine (5th ed., pp. 585-594). Philadelphia: Elsevier Saunders.
 Go back to the Dream Library index. Go back to the Dream Library index.
|
