The Relationship Between Dreaming and Autonoetic Consciousness: The Neurocognitive Theory of Dreaming Gains in Explanatory Power by Drawing Upon the Multistate Hierarchical Model of Consciousness
G. William Domhoff
University of California, Santa Cruz
|
NOTE: If you use this paper in research, please use the following citation, as this on-line version is simply a reprint of the original article:
|
|
Domhoff, G. W. (2023). The relationship between dreaming and autonoetic consciousness: The neurocognitive theory of dreaming gains in explanatory power by drawing upon the multistate hierarchical model of consciousness. Dreaming, 33(1), 1-18.
|
|
[Download PDF of this article]
|
Abstract
The neurocognitive theory of dreaming, which emphasizes portions of the default network as the most important neural substrates that support dreaming, can increase its explanatory power by drawing upon the multistate hierarchical model of consciousness (Domhoff, 2022; LeDoux, 2019). The default network's two main subsystems, the dorsal medial prefrontal cortex subsystem and the medial temporal cortex subsystem, are involved in supporting imagination and mind-wandering during waking, which suggests that dreaming is a form of spontaneous imaginative thought. The multistate hierarchical model of consciousness considers the regions that support dreaming to be part of the intermediate multimodal areas in the multistate hierarchy. In terms of self-reflective (autonoetic) consciousness, the schemas developed and stored in the intermediate multimodal areas are re-represented by the higher-order networks in the anterior prefrontal cortex (the dorsal lateral and ventral lateral prefrontal cortices, and the lateral frontal pole), which are the necessary additional regions essential for autonoetic consciousness to emerge. These anterior areas are relatively deactivated during all stages of sleep, which may help explain various "cognitive insufficiencies" during dreaming, such as the lack of autonoetic consciousness and episodic memory, and the relative absence of figurative thinking and emotions. Dreaming produces only noetic (fact-knowing) consciousness, based on semantic memory. However, there are rare instances of self-awareness during dreaming, in which the anterior higher-order networks are atypically activated during sleep. The findings on the immaturity of the default network until ages 9-11, and the gradual development of the cognitive abilities necessary for dreaming, are consistent with this analysis.
Introduction
This proposed extension of the theoretical understanding of the relationship between dreaming and self-reflective (autonoetic) consciousness begins with the claim that two relatively new and recently updated theories, the neurocognitive theory of dreaming and the multistate hierarchical model of consciousness, are highly compatible (Domhoff, 2022; LeDoux, 2019; LeDoux & Lau, 2020). More specifically, the explanatory power of the neurocognitive theory of dreaming therefore can be increased by drawing upon the more encompassing and more fully delineated multistate hierarchical model of consciousness, which was developed without any focus on dreaming. Most importantly, the multistate hierarchal model can explain dreams as a facet of its intermediate multimodal areas, which includes many of the same neural substrates that are claimed to be the neural substrate for dreaming in the neurocognitive theory of dreaming. The multistate hierarchal model therefore makes it possible to explain the several differences between dreaming and autonoetic consciousness — such as the lack of episodic memories during dreaming and the relative absence of figurative thinking and emotions — as "cognitive insufficiencies" during dreaming. The concept of cognitive insufficiencies encompasses (1) cognitive defects due to brain lesions; (2) temporary deficiencies due to immaturity; and (3) recurring deficiencies during sleep due to the relative deactivation of neural substrates that are activated during waking, as discussed in the next section (Domhoff, 2022, pp. 55-56,160-161, 234-235, for discussions of cognitive insufficiencies during dreaming).
Drawing upon the multistate hierarchical model of consciousness and the work of other neurocognitive researchers, it also may be possible for the neurocognitive theory of dreaming to explain the rare phenomenon of self-awareness during dreaming. It does so in terms of the atypical activation during sleep of the most anterior areas of the higher-order network (the dorsal lateral and ventral lateral prefrontal cortices, and the lateral frontal pole), which are the necessary additional regions essential for autonoetic consciousness to emerge according to the multistate model (LeDoux, 2019, pp. 359-362; 2021, p. R828, Figure 3). (The lateral frontal pole, which is the most recent and anterior portion of the human brain, is called the "rostrolateral prefrontal cortex" and the "lateral frontopolar cortex" by the other neurocognitive researchers who will be drawn upon in this article (e.g., Christoff & Gabrieli, 2000; Dixon, Fox, & Christoff, 2014a).)
More generally, the discussion of these possibilities also may serve to stimulate thinking about the compatibility of various theories of dreaming and theories of autonoetic consciousness. Perhaps, for example, the neurocognitive theory of dreaming may be compatible with other theories of autonoetic consciousness, or other theories of dreaming may be compatible with the multistate hierarchical model of consciousness. And, of course, other theories of dreaming may be compatible with various theories of autonoetic consciousness, including several contemporary theories of consciousness that also discuss dreaming on the basis of empirical dream research. However, a discussion of these other contemporary theories — all of which differ from each other, and from the neurocognitive theory of dreaming and the multistate hierarchical model of consciousness — is beyond the purview of this narrowly focused article.
It also is necessary to emphasize what this article is not claiming. The assertion that the neurocognitive theory of dreaming is compatible with, and therefore can draw upon, the multistate hierarchical model of consciousness does not imply that either of these theories is correct. Compatibility is one issue, the adequacy of the two theories is another. Nor does the compatibility discussed in this article imply the other theories that focus on the relationship between dreaming and autonoetic consciousness are incomplete or wrong.
The Neurocognitive Foundations of Dreaming
Neuroimaging studies of sleep reveal that the neural substrates that subserve four of the five association networks active during waking thought — the frontoparietal control network, the dorsal attention network, the salience/ventral network, and the limbic network — are relatively deactivated during sleep, as are the neural substrates that subserve the visual and sensorimotor networks (Braun et al., 1997; Braun et al., 1998; Dang-Vu, Schabus, Desseilles, Schwartz, & Maquet, 2007; Fox, Nijeboer, Solomonova, Domhoff, & Christoff, 2013; Maquet et al., 1996; Nofzinger, Mintun, Wiseman, Kupfer, & Moore, 1997; Uitermarkt, Bruss, Hwang, & Boes, 2020). The exception is the relative activation of portions of the default network, the fifth of the association networks. More precisely, the dorsal medial prefrontal cortex subsystem and the medial temporal cortex subsystem of the default network are activated during the sleep-onset process, NREM 2 sleep, and REM sleep (Domhoff, 2011; Eichenlaub, Bertrand, & Ruby, 2014; Sämann et al., 2011; Stevner et al., 2019; Tagliazucchi et al., 2013; Tarun et al., 2021).
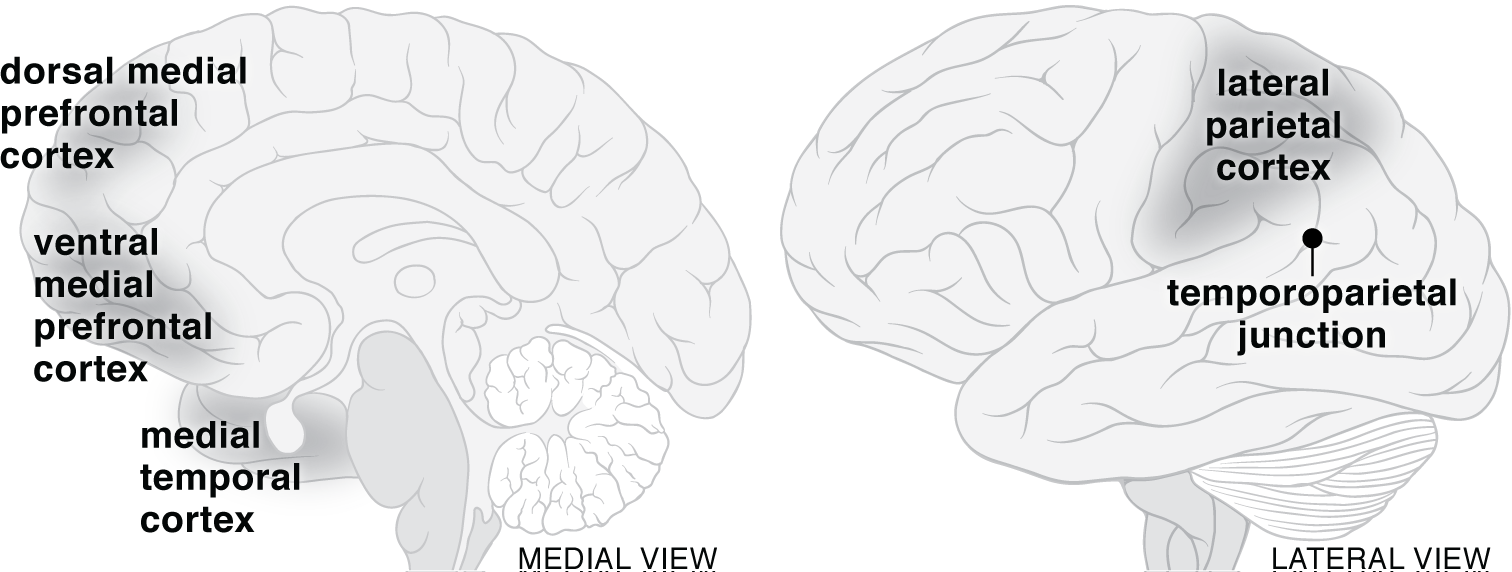
These two subsystems, which are the primary neural basis for dreaming, are also involved in supporting imagination and mind-wandering in waking life when the frontoparietal control network, the dorsal attention network, and the salience/ventral network are not constraining the default network to the degree they do in task-oriented contexts (Andrews-Hanna, Reidler, Sepulcre, Poulin, & Buckner, 2010; Andrews-Hanna, Smallwood, & Spreng, 2014; Chang, Liu, Chen, Liu, & Duyn, 2013; Christoff, Irving, Fox, Spreng, & Andrews-Hanna, 2016; Fox, Spreng, Ellamil, Andrews-Hanna, & Christoff, 2015; Spreng, Madore, & Schacter, 2018). Although the two subsystems of the default network are relatively activated during dreaming, the "zones of integration" (the areas in the default network that connect it to the other association networks during waking), are relatively deactivated throughout sleep. Their relative deactivation contributes to the independence of the default network at the times when it is relatively activated during sleep. Figure 1 provides an approximate indication of the locations of the main regions in the default network that are activated during dreaming. However, it does not include all of the neural substrates that are activated during dreaming. A more detailed presentation of the brain areas that support dreaming is provided in Table 1 at the end of the next section, which shows the differences between the neural networks that support autonoetic consciousness and dreaming.
During the sleep-onset process, NREM 2 sleep, and REM sleep, the dorsal medial and medial temporal subsystems of the default network are augmented by the relative activation of secondary sensory cortices (including the lingual gyrus), along with secondary sensorimotor areas and the caudate nucleus, which combine to create the neural substrate that subserves dreaming (Domhoff, 2022, Chapter 2). However, the truncated nature of this neural substrate leads to the lack of several cognitive capabilities during dreaming that are enabled during waking by interactions among two or more of the five association networks: an awareness of the here and now, the ability to access episodic memories (Baylor & Cavallero, 2001; Fosse, Hobson, & Stickgold, 2003; Malinowski & Horton, 2014), the near or complete absence of figurative thought and symbolism (Domhoff, 2022, Chapter 5), and the relatively infrequent inclusion of emotions (Domhoff, 2022, Chapter 8). As noted above, dreaming therefore suffers from several cognitive insufficiencies when compared to the cognitive capabilities that are generally available during waking thought.
The presentation of a full statement concerning the cognitive processes supported by the neural substrates that are active during dreaming is outside the narrowly defined purpose of this theoretical article. However, it has to be added that dreaming is a form of "simulation," which is defined as "a particular type or subset of thinking that involves imaginatively placing oneself in a hypothetical scenario and exploring possible outcomes" (Schacter, Addis, & Buckner, 2008, p. 42). Since simulation during dreaming also involves vivid mental imagery, due to the relative activation of secondary sensory and sensorimotor cortices, the enhanced simulation that enables dreaming can be characterized as "embodied simulation," which is defined in the strict psychological sense of this term as off-line cognition that is body based. More specifically, embodied simulation occurs when "sensory and motor resources are brought to bear on mental tasks whose referents are distant in time and space or are altogether imaginary" (Wilson, 2002, p. 635, italics added).
Within that definition, embodied cognition includes mental imagery, imagination, figurative thinking, episodic memory, reasoning, and problem-solving, and there is ample evidence for its occurrence during waking states (e.g., Bergen, 2012; Calvo-Merino, Jones, Haggard, & Bettina, 2017; Gibbs, 2006; Landau, Meier, & Keefer, 2010; MacDonald & Culham, 2015; Mason & Just, 2020; Wilson, 2002). It is within this framework, and the evidence for it, that dreaming can be understood as an intensified and enhanced form of mind-wandering and daydreaming, which occurs whenever the frontoparietal control network, the dorsal attention network, and the salience/ventral network are not constraining the default network to the extent that they do in task-oriented contexts during alert waking (Andrews-Hanna et al., 2010; Andrews-Hanna et al., 2014; Chang et al., 2013; Christoff, Gordon, Smallwood, Smith, & Schooler, 2009; Christoff et al., 2016; Fox et al., 2015).
The emphasis on portions of the default network as the primary basis for dreaming during the sleep-onset process, NREM 2 sleep, and REM sleep is supported by studies of the impact of brain lesions on dreaming (Bischof & Basset, 2004; Jus et al., 1973; Kerr, Foulkes, & Jurkovic, 1978; Poza & Marti-Masso, 2006; Solms, 1997). These studies make use of patients with focal lesions, most of whom were studied before neural plasticity could lead to compensations for the lesions. They can be thought of as a series of inadvertent, experimentally-based tests of the neuroimaging findings. Studies of the impact of lesions in many different areas in the brain show that patients can lose dreaming due to lesions in either the medial prefrontal cortex or the temporoparietal junction, which are parts of the default network. Patients also report specific visual changes after lesions in various areas in the secondary visual cortex. On the other hand, lesions in the dorsal lateral prefrontal cortex, amygdala, primary visual cortex, primary sensorimotor cortices, hypothalamus, cerebellum, and several areas in the brainstem have no impact upon dreaming (Blake, Terburg, Balchin, van Honk, & Solms, 2019; Solms, 1997, pp. 82, 153-154, 219-223, 237). The following generalization therefore can be made on the basis of a large number of cases: lesions outside the neural substrates that subserve dreaming have no impact on dreaming, but lesions within the neural substrates that subserve dreaming have an effect on dreaming, ranging from the complete loss to dreaming to the loss of different visual aspects of dreaming (Domhoff, 2022, pp. 28-33, for the evidence for this generalization).
In combination, the neuroimaging and lesion studies circumscribe the boundaries of the neural substrates necessary for dreaming. They provide a form of convergent validity for the claim that the general contours of the neural substrate that subserves dreaming are now fairly well established, although much still remains to be learned about its inner workings. The lesion studies add first-person subjective accounts concerning the presence or absence of dreaming, and in the process eliminate many neural substrates as possible areas that might subserve dreaming. More generally, the neurocognitive theory of dreaming, by drawing upon both neuroimaging studies and lesion studies, claims that dreaming is a property of a specific neurocognitive network. If that network is not intact, dreaming is defective or does not occur at all.
In addition, the theory has a developmental dimension and can assimilate the many findings on dream content, which have been developed on the basis of quantitative content analyses of many thousands of dream reports collected from both children and adults inside and outside of sleep-dream laboratories (Domhoff, 1996; 2022, Chapters 3-4 and 6-8). The developmental dimension is briefly overviewed in a later section of this article because findings on the gradual maturation of the default network, and the parallel development of dreaming and the waking cognitive capacities that are very likely necessary for dreaming, provide further support for the analysis based on adult participants in this and the following section. The findings on dream content, on the other hand, are not relevant to the focus of this article.
The Multistate Hierarchical Model of Consciousness
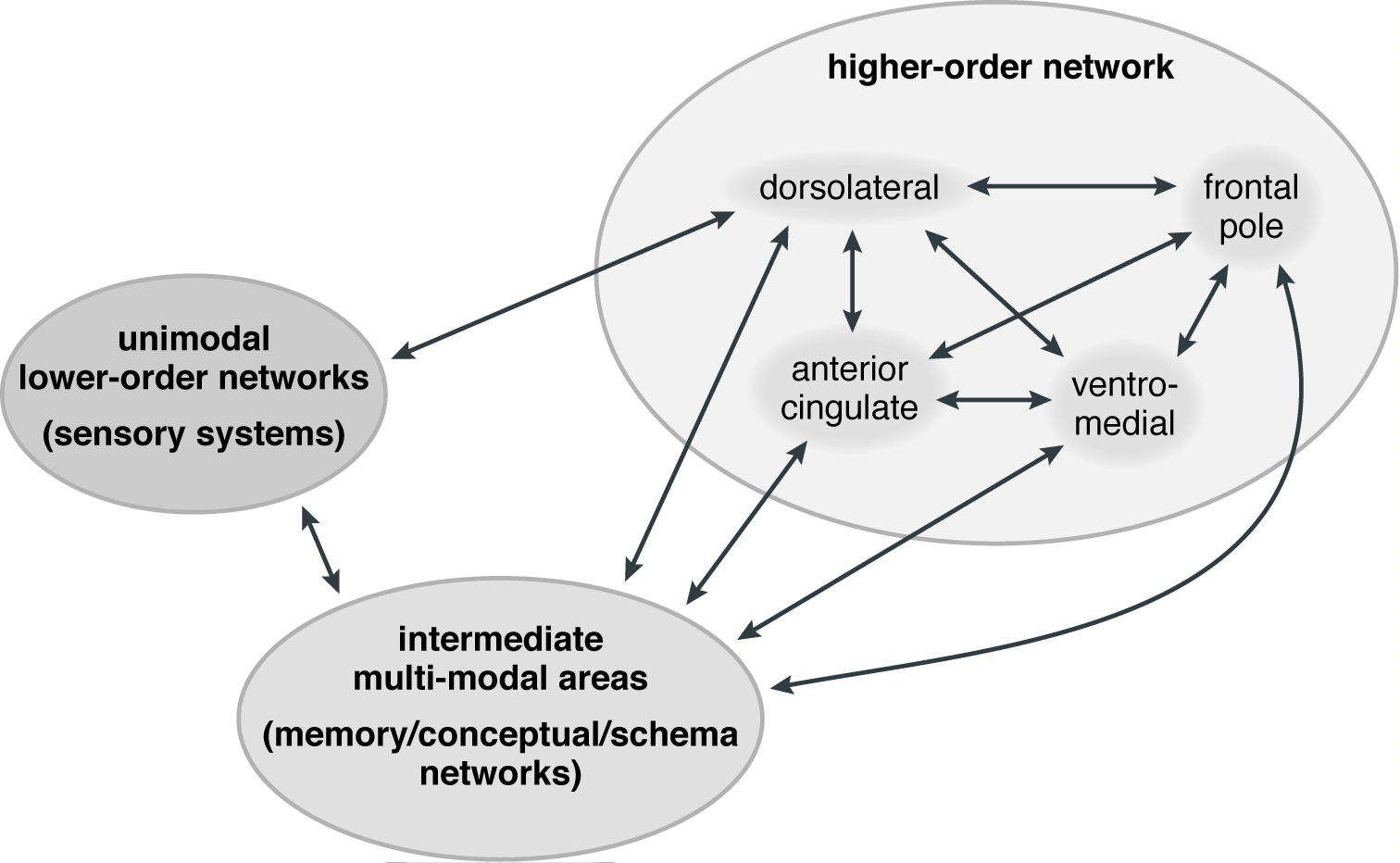
The multistate hierarchical model of consciousness is based on a concept called "cortical consciousness networks." This concept has parallels with the concept of "association networks" that is used in the neurocognitive theory of dreaming. From the perspective of the multistate hierarchical model, several of the areas that support dreaming can be understood as intermediate multimodal areas in the hierarchy that leads to autonoetic consciousness (LeDoux, 2019, pp. 282-283, 306-308). In waking thought, these intermediate multimodal areas first all of have an important role in processing all forms of incoming sensory information from the unimodal lower-order networks and assimilating them into existing memory banks. The networks within the intermediate multimodal areas also are crucial in creating "schemas," which are "complex collections of memories about specific things, such as situations, one's self, and also about emotions" (LeDoux, 2021, p. R828). These areas, which include the ventral medial and dorsal medial cortices that are essential to dreaming, are thus "sources of complex cognitive representations that can, like perceptual, mnemonic, and conceptual representations, be used in the assembly of conscious experiences by the higher-order network" (LeDoux, 2019, p. 360).
The intermediate multimodal areas then connect to the higher-order network located in the most anterior portions of the prefrontal cortex. These anterior areas include the dorsal and ventral lateral prefrontal cortices and the lateral frontal pole, the latter of which is unique in its size and patterns of connectivity to the cognitively modern humans that emerged between 50,000 and 200,000 years ago (LeDoux, 2019, pp. 359-362, 373; 2021, p. R828, Figure 3). However, due to the far greater connectivity between prefrontal and parietal areas in the human brain than in other mammals, including other primates, the parietal cortex also contributes to the executive functions carried out by the higher-order network (LeDoux, 2019, pp. 258-259; 2020, pp. R2, R3). Within this context, the re-representation of the information from the intermediate multimodal networks within the higher-order network is the additional step that makes autonoetic, self-reflective consciousness possible. The lateral frontal pole, which "only receives input from the multimodal convergence zones, and creates the most abstract conceptual presentations in the brain," works closely with the dorsal and ventral lateral prefrontal regions in coordinating top-down control of all aspects of human thinking and action (LeDoux, 2019, pp. 252-253). Put another way, these areas "together allow executive control over both unimodal (sensory) and multimodal (conceptual) processing in posterior areas, as well as control of deliberative behavior by way of connections to the motor cortex" (LeDoux, 2019, p. 253). A schematic graphic of the three levels of the theory is presented in Figure 2, in which the two-way arrows emphasize both the complexity of the interactions among the three levels and the fact that the higher-order network has direct pathways through which it can control the unimodal lower-order networks, as well as the multimodal memory/conceptual/schematic networks.
Due to the importance of the most anterior regions of the prefrontal cortex in supporting autonoetic consciousness, both the multistate hierarchical model of consciousness and the neurocognitive theory of dreaming imply that dreaming has different properties than does autonoetic consciousness. Unlike during waking, the higher order network is deactivated according to the multistate model, and four of the five association networks are relatively deactivated during dreaming according to the neurocognitive theory of dreaming. In addition, the primary sensory and sensorimotor networks are relatively deactivated, and incoming stimuli are gated in the thalamus (Chow, Horovitz, Picchioni, Balkin, & Braun, 2013; Picchioni et al., 2014; Tarun et al., 2021). Unlike autonoetic consciousness, dreaming therefore does not have the perceptual inputs that represent the demands and opportunities present in the waking world, which are sent to relevant memory banks for recognition and incorporated into schemas. It also lacks the capacity to redirect thought through top-down commands — a capacity supported by the lateral frontopolar, dorsal lateral and ventral lateral prefrontal regions (LeDoux, 2019, pp. 359-362). This emphasis on top-down commands is paralleled and supported by the findings on "cognitive control networks" by those theorists who work within the framework provided by the frontoparietal control network and the four other association networks (e.g., Christoff & Gabrieli, 2000; Christoff et al., 2016; Christoff et al., 2001; Dixon et al., 2014a; Dixon, Fox, & Christoff, 2014b). By cognitive control these theorists mean "the process by which PFC [i.e., prefrontal cortex] selectively biases currently relevant representations in other parts of the brain, thereby helping focus attention on currently relevant stimuli while diminishing attention toward other competing stimuli" (Christoff, 2014, p. 319).
Since dreaming is entirely self-generated and receives no external feedback, it depends on the semantic memories and the schemas at the intermediate multimodal level to generate imaginative embodied simulations. These embodied simulations often (but not always) focus on the concerns of the self-system, which is enabled in large measure by regions within the default network (Abraham, 2013; D'Argembeau, 2020; Domhoff, 2022, Chapters 2-3).To provide a contrast with daydreaming — the spontaneous mental state that appears to be most similar to dreaming — people can "realize" (reflect upon) the fact that they are daydreaming (Andrews-Hanna et al., 2014; Christoff et al., 2016; Fox, Andrews-Hanna, & Christoff, 2016). A somewhat simplified comparison of the brain networks that subserve autonoetic consciousness during waking, and dreaming during NREM 2 sleep, and REM sleep, is provided in Table 1.
Table 1. A comparison of the main neural substrates that support autonoetic consciousness and dreaming.
| Brain Region | Consciousness Network | Dreaming Network |
|---|
| Frontal pole/Rostrolateral PFC | activated | relatively deactivated |
| Dorsolateral PFC | activated | relatively deactivated |
| Ventrolateral PFC | activated | relatively deactivated |
| Orbital PFC | activated | relatively deactivated |
| Ventromedial PFC | activated | activated |
| Dorsomedial PFC | activated | activated |
| Episodic memory network | activated | relatively deactivated |
| Semantic memory network | activated | activated |
| Lingual gyrus (visual processing) | activated | activated |
| Secondary visual areas | activated | activated |
| Primary visual cortex | activated | relatively deactivated |
NOTE: This table is based on the discussions and graphics in Domhoff (2022, Chapter 2, with special reference to Table 2.2), and in LeDoux (2019, Chapter 57, especially Figures 57.1 and 57.2).
Self-Awareness During Dreaming in a Hybrid Sleep State
As one team of neurocognitive researchers notes, "humans typically lack awareness that they are dreaming while dreaming" and "most individuals experience lucid dreams [dreams during which self-awareness occurs] rarely" (Baird, Castelnovo, Gosseries, & Tononi, 2018, p. 1). Nevertheless, self-awareness during dreaming is of interest in terms of this theoretical article because it is a dream state, however rare, that includes self-awareness. This self-awareness occurs occasionally for some people after returning to bed following an early-morning trip to the bathroom, or shortly before their spontaneous morning awakening, which suggests that the brain may be more generally activated when self-awareness during dreaming occurs. But little is known about those who experience self-awareness during dreaming more frequently, or why they do so. Some reports by or about frequent "lucid dreamers," as they call themselves, suggest that self-awareness during dreaming may begin because of frightening night experiences as a child, which make falling asleep anxiety-arousing to think about, and thereby cause sleep to be fitful. For others, self-awareness during dreaming begins while dealing with insomnia, or with the discontinuance of the ingestion of psychedelic drugs (Lucidity Newsletter, 1986; Rolston, 2013).
According to one systematic waking-state fMRI neuroimaging study, there may be neurological differences between those who do and do not report having experienced self-awareness during dreams with a high frequency. These differences could be caused by one or more of several factors, including genetic predispositions, the development of atypical cognitive capacities, or some of the events mentioned in the previous paragraph. The study compared 14 participants who reported they had 3 or more instances of self-aware dreaming per week with a control group of similar size, whose members reported they had 1 or fewer self-awareness dreams per year. The researchers concluded that self-awareness during dreaming may be "associated with increased functional connectivity between aPFC [the anterior prefrontal cortex] and temporoparietal association areas, regions normally deactivated during sleep" (Baird et al., 2018, p. 1). This finding is consistent with the emphasis in the neurocognitive theory of dreaming on the relative deactivation of these areas during normative dreaming.
Two sleep-lab studies yielded results similar to those in the waking-state study. The first study, which used high-density EEG, involved 6 highly motivated college students who remembered 3 or more self-aware dreams each night at home. The participants each spent up to 5 nights in the sleep lab; 3 of them were able to signal 1 instance each of self-awareness during dreaming (Voss, Holzmann, Tuin, & Hobson, 2009, p. 1191). Their brain patterns were characterized by "wake-like inter-scalp networking, including high-frequency bands," which were "most pronounced in frontal and frontolateral coherences;" the authors concluded that self-awareness during dreaming may occur during a "hybrid state" that has features of waking as well as REM sleep (Voss et al., 2009, p. 1196). These findings support the emphasis on portions of the default network as the basis for imagination and dreaming in the neurocognitive theory of dreaming, and the emphasis on the dorsal lateral prefrontal cortex, the ventral lateral prefrontal cortex, and the lateral frontal pole as the additional necessary ingredients for autonoetic consciousness in the multistate hierarchical model of consciousness.
A nighttime combined EEG/fMRI study led to similar results. It was based on 15 nights of fMRI studies that made use of 4 adult male participants between the ages of 27 and 31, who had actively tried to have greater self-awareness during dreaming for several years or more for their own reasons. Although numerous REM periods occurred over the space of the 15 nights, only 2 of the 4 participants were able to provide eye-movement signals in 1 instance each, which were corroborated by their post-awakening reports. These results reinforce the finding that lucid dreaming is rare even for those who strive to experience it. However, the two occurrences seemed clear enough to report "a reactivation of several areas normally deactivated during REM sleep," and especially the dorsal lateral prefrontal cortex, ventral lateral prefrontal cortex, and lateral frontopolar cortex (Dresler et al., 2012, p. 1020). These areas are part of the higher-order network in the multistate hierarchical model of consciousness. (Here it may be useful to recall that "lateral frontopolar cortex," "rostrolateral prefrontal cortex," and "lateral frontal pole" are three different names for the same brain area, which makes it clear that many neurocognitive researchers agree on the important role this area plays in self-reflective awareness.)
It is also relevant in terms of the emphasis on the imaginative nature of dreams in the neurocognitive theory of dreaming that dream reports collected from individuals who report self-awareness during dreaming do not differ from other dream reports. In a comparison of 441 lucid and non-lucid dream reports collected from 10 different samples of college students and older adults over the space of several years, relatively few differences were found. There were more auditory and kinesthetic elements in self-aware dreams, and fewer characters and less happiness in the non-self-aware dreams, but the social interactions were similar, leading to the conclusion that the dream reports from the two different neurocognitive states "are more alike than they are different" in terms of their content (Gackenbach, 1988, pp. 192-193, italics in the original). In addition, as a comprehensive overview of the relevant literature concluded, "a majority of lucid dreams, it seems, feel very much like standard wakefulness and nonlucid dreams" (Windt, 2015, p. 118).
Based on this overview of the portions of the literature on self-awareness during dreaming that are relevant to a neurocognitive explanation for this rare phenomenon, the imaginative nature of such dreams can be explained by the relative activation of portions of the default network, secondary sensory areas, and the caudate nucleus, which provide the neural foundations for the neurocognitive theory of dreaming. The presence of self-awareness may be explained by the atypical relative activation of the dorsal lateral prefrontal cortex, the ventral lateral prefrontal cortex, and lateral frontal pole (Dresler et al., 2012, p. 1020; Voss et al., 2009), which are the additional necessary ingredients for self-awareness in the multistate hierarchical model of consciousness. However, these results need to be replicated with larger sample sizes, and the factors that allow self-awareness to be possible during dreaming without awakening remain unexplained. Put another way, the extent to which self-awareness during dreaming is similar to autonoetic consciousness during waking remains to be fully examined.
One possibility might be that a specific network of areas in the anterior prefrontal cortex, including the lateral frontal pole/rostrolateral prefrontal cortex, serve to focus attention internally or externally (e.g., Burgess, Dumontheil, & Gilbert, 2007; Christoff, 2014; Christoff & Gabrieli, 2000; Christoff, Ream, Geddes, & Gabrieli, 2003; Henseler, Krüger, Dechent, & Gruber, 2011). In particular, a study showing that there are two subsystems within the frontoparietal control network, one that regulates the dorsal attention network during attention and action, and one that interacts with the default network when the focus is on personal thoughts, might provide a useful starting point (Dixon et al., 2018). Two other studies support the possibility that the internally oriented portion of the frontoparietal control network may play a role in self-awareness during dreaming. First, a waking study of the lateral frontal pole/rostrolateral prefrontal cortex found that participants could gain some control of self-referential thoughts relating to that region by reflecting on their own thoughts ("meta-awareness") (McCaig, Dixon, Keramatian, Liua, & Christoff, 2011). In addition, still another waking study found that this same internally oriented region within the frontoparietal control network interacts with regions in the default network that are concerned with self-referential information and contribute to meta-awareness (Dixon et al., 2022).
Parallel Developmental Paths Support Dreaming and Autonoetic Consciousness
The findings described in the earlier sections of this article concerning the basis of normative everyday dreaming and autonoetic consciousness are consistent with what has been learned about the parallel development of dreaming and autonoetic consciousness in young children. The most important finding from longitudinal and cross-sectional developmental studies of dreaming is that dreaming does not occur often, if at all, before age 5, and develops only gradually in terms of its frequency and complexity between the ages of 5- 9. Dreaming then becomes somewhat more adultlike between ages 9-11 but dream content does not become similar to that of adults until ages 12-15 (Domhoff, 2022, Chapters 6-7, for a synthesis; Foulkes, 1982, 2017; Foulkes, Hollifield, Sullivan, Bradley, & Terry, 1990; Strauch, 2004, 2005; Strauch & Lederbogen, 1999, for the longitudinal and cross-sectional studies that provide the basis for this synthesis). In addition, neuroimaging studies of children and adolescents reveal that the default network matures only slowly, and it is not close to adultlike levels until ages 9-13, which is consistent with the emphasis in the neurocognitive theory of dreaming on the role of portions of the default network (Domhoff, 2022, Chapter 6, especially Figure 6.1; Fair et al., 2008; Fair et al., 2009; He et al., 2019; Moraczewski, Nketia, & Redcay, 2020; Sherman et al., 2014).
Moreover, dreaming seems to require the development of five cognitive capacities that have been systematically studied by those psychological scientists who focus on waking cognitive development. These cognitive capacities include the ability to form concepts, generate mental imagery, make use of narrative thinking, and think imaginatively, along with the acquisition of an autobiographical self. It is likely that these are the cognitive abilities that are necessary for the more general cognitive process of simulation, as will become clear as this section unfolds. The ability to form concepts develops between late infancy and 30 months (Mandler, 2008, 2012; Nelson, 2004). Children gradually develop the capacity to generate mental imagery, make use of narrative thinking, and think imaginatively between the ages of 3 and 6, but imaginative abilities only gradually become more adultlike until ages 10-12 (Coughlin, 2016, p. 22; Coughlin, Robins, & Ghetti, 2019; Foulkes et al., 1990; Nelson, 2007, pp. 170-171; Taylor, 2013). The many findings in this large waking-state literature in developmental psychology have been summarized and synthesized in the context of adding a developmental dimension to the neurocognitive theory of dreaming (Domhoff, 2018, Chapter 4; 2022, Chapters 6-7).
With regard to the development of autonoetic consciousness, it is first of all relevant that preschool children "don't understand that your thoughts can be internally generated," or that "thoughts can simply follow the logic of your internal experiences instead of being triggered from the outside," until the ages of 5-6 (Gopnik, 2009, p. 152). Similarly, pre-school children do not understand that dreams are internally generated, and they sometimes think dreams are a shared experience, which may be because they do not dream very often and therefore rely on what parents and children's books tell them about dreaming (Meyer & Shore, 2001; Woolley & Boerger, 2002; Woolley & Wellman, 1992). In addition, young children "don't experience their lives as a single timeline stretching back into the past and forward into the future;" nor do they "feel immersed in a constant stream of changing thoughts and feelings" (Gopnik, 2009, p. 153). Furthermore, if pre-school children are asked during an idle moment what they were just thinking, they usually say "nothing," which actually may be the case if they are not proficient at imagining and simulating. In a developmental study of children's familiarity with mind-wandering, only 44% of children understood what mind-wandering is at ages 6-7, compared to 86% by ages 10-11. This finding raises the possibility that mind-wandering has a developmental trajectory similar to that of dreaming, which again suggests the importance of a mature default network and the cognitive ability to simulate in order to mind-wander and dream (Eisbach, 2013, Table 2).
Based on these and other findings, developmental psychologist Allison Gopnik (2009, p. 153) argues that children need to have an autobiographical self in order to experience autonoetic consciousness, which according to her analysis does not develop until about age 5 and thereafter. Developmental psychologist Katherine Nelson and her colleagues have enriched the understanding of an autobiographical self through numerous studies, and report a similar developmental timeline. Their work demonstrates that there is an important narrative dimension to autonoetic consciousness, which develops through pre-school children's conversations with adults, especially their mothers, about recent past events and upcoming events (Nelson, 2005; Nelson & Fivush, 2020).
Memory researcher Endel Tulving (2005) reaches a similar conclusion about the importance of autobiographical memories in the development of autonoetic consciousness after assessing the literature on the development of episodic memory in children, which manifests itself between ages 4 and 5. In his view, autonoetic consciousness appears to be a necessary correlate of episodic memory, which includes a sense of subjective experience, a sense of inner existence, and a personal identity that persists and grows over time (Tulving, 2005, pp 29-34). This pioneering analysis of the relationship between episodic memory and autonoesis is also considered to be essential in the multistate hierarchical model of consciousness (LeDoux, 2019, pp. 294-299; LeDoux & Lau, 2020, p. R109).
The emphasis on the importance of autobiographical memory in the development of autonoetic consciousness has a parallel with the development of dreaming. The few and brief accounts of dreams by preschool children rarely include the dreamer herself or himself until ages 5-6. The presence and involvement of the dreamer in the ongoing dream is still not yet fully present in dream reports at ages 7-8, which suggests that both noetic dreaming and autonoetic consciousness gradually develop between ages 5 and 8 (Foulkes, 1999, pp. 146-150, for an account of consciousness as a developmental construction; 2017, pp. 4-5, 12, for an updated view of consciousness that draws upon Tulving's work on episodic memory and LeDoux's work on emotion; LeDoux, 2015). The overall findings in this section therefore provide further support for a conclusion drawn earlier in this article: autonoetic consciousness during waking, and dreaming during sleep onset, NREM 2 sleep, and REM sleep, have a close relationship, but they do not have quite the same properties.
Implications and Conclusions
Considering the adult and developmental findings together, the neurocognitive theory of dreaming is compatible with the multistate hierarchical model of consciousness. The multistate hierarchical model therefore can be drawn upon to augment the explanatory power of the neurocognitive theory of dreaming. From the perspective of the neurocognitive theory of dreaming, dreaming occurs during the sleep-onset process, NREM 2 sleep, and REM sleep due to the concomitant relative activation of portions of the default network and secondary sensory and sensorimotor cortices, along with the caudate nucleus. The combination of spontaneous imaginative thought, vivid mental imagery, and the sense of movement that sometimes occurs during drifting waking thought and daydreaming are transformed into an enhanced version of embodied simulation during dreaming (Domhoff & Fox, 2015; Fox et al., 2013).
Dreaming has different properties than autonoetic consciousness does because the neural substrates that support dreaming lack the perceptual inputs that serve as guideposts for thinking during waking life, as well as the top-down capacity to redirect thought. Dreaming also lacks the neural support that enables access to episodic memory and figurative thinking, and includes emotion much less often than is generally assumed (Domhoff, 2022, Chapter 8). These cognitive insufficiencies are very likely due to the relative deactivation of the frontoparietal control network, the attention networks, and the limbic network according to the neurocognitive theory of dreaming, and to the relative deactivation of the higher-order network according to the multistate hierarchical model of consciousness. The relative deactivation of these particular networks during dreaming means that autonoetic consciousness cannot be supported according to both the neurocognitive theory of dreaming and the multistate hierarchical model of consciousness. However, it is likely that the generally rare episodes of self-awareness during dreaming involve the relative activation of internally oriented portions of the anterior prefrontal cortex during an atypical (hybrid) sleep state (e.g., Dixon et al., 2022; Dixon et al., 2018; McCaig et al., 2011). This possibility needs to be explored in more detail in studies with larger sample sizes of self-identified lucid dreamers that include neurocognitive assessments of the participants as well as detailed life histories.
The immaturity of the default network and the gradual development of cognitive capacities until ages 9-11, including the ability to simulate, join together to explain the infrequency of dreaming and the lack of complexity and self-involvement in children's dream reports until that age period. In addition, the parallels in the development of the cognitive capacities that lead to noetic dreaming during sleep onset, NREM 2, and REM sleep, as well as autonoetic consciousness during waking, provide further evidence that dreaming and autonoetic consciousness have a close relationship, even though dreaming has different properties than does autonoetic consciousness. From the perspective of the neurocognitive theory of dreaming, both dreaming and autonoetic consciousness are the properties of specific networks. If the networks are immature or damaged in certain areas, then dreaming or autonoetic consciousness will be defective in some way or will not occur at all.
The areas that support the noetic mental state of dreaming are important as part of the intermediate multimodal areas in the multistate hierarchical model of consciousness. In that model, the memories, concepts, and schemas from the intermediate multimodal areas are re-represented as inputs into the higher-order network, and thereby contribute to autonoetic consciousness during waking (LeDoux, 2019, pp. 359-361). In this view, the additional necessary step that leads to autonoetic consciousness is supported by brain areas anterior to the default network — namely, the dorsal lateral and ventral lateral regions of the prefrontal cortex, and the lateral frontal pole. Dreaming therefore has different properties than does autonoetic consciousness within the context of both the multistate hierarchical model of consciousness, with its emphasis on intermediate multimodal areas that produce schemas, and the neurocognitive theory of dreaming, with its emphasis on portions of the default network and the cognitive capacities those portions are able to support.
In closing, it may be useful to mention again what this article is not claiming. It is not a brief for the accuracy or usefulness of either of the two theories because it has not discussed the adequacy of the evidence that supports them, a task that has been carried out by the individual theorists in their presentations of their respective theories (Domhoff, 2020a, 2022; LeDoux, 2019, 2020; LeDoux & Lau, 2020; LeDoux, Michel, & Lau, 2020). Nor has the article anticipated and attempted to answer the likely criticisms and claims by proponents of other views, which is once again an undertaking that has been attempted in other venues (Domhoff, 2020b; 2022, Chapters 8-11; LeDoux, 2019, 2020, 2021; LeDoux & Lau, 2020). Instead, this article has focused on the increased explanatory power provided to the neurocognitive theory of dreaming by drawing upon the multistate hierarchical model of consciousness.
Acknowledgements
Many thanks to the reviewers for insisting that the analysis presented in this article should be presented more fully and with more detail than it was originally. Thanks, also, to Matthew L. Dixon for sharing his invaluable expertise and suggesting references concerning the frontal pole and the frontoparietal control network, and to Joseph LeDoux for reading the section on the multistate hierarchical theory of consciousness and for suggesting references on the frontal pole. Further thanks to Matthew Erdelyi for suggesting that graphics of the relevant brain networks might be helpful, and to my research assistant and frequent co-author, Adam Schneider, for making the graphics and suggesting many helpful clarifications and word changes in the text.
References
Abraham, A. (2013). The world according to me: personal relevance and the medial prefrontal cortex. Frontiers in Human Neuroscience, 7, 341-344.
Andrews-Hanna, J., Reidler, J., Sepulcre, J., Poulin, R., & Buckner, R. (2010). Functional-anatomic fractionation of the brain's default network Neuron, 65, 550-562.
Andrews-Hanna, J., Smallwood, J., & Spreng, R. N. (2014). The default network and self-generated thought: component processes, dynamic control, and clinical relevance. Annals of the New York Academy of Science, 1316, 29-52.
Baird, B., Castelnovo, A., Gosseries, O., & Tononi, G. (2018). Frequent lucid dreaming associated with increased functional connectivity between frontopolar cortex and temporoparietal association areas. Science Reports, 8(17798). doi:10.1038/s41598-018-36190-w
Baylor, G., & Cavallero, C. (2001). Memory sources associated with REM and NREM dream reports throughout the night: A new look at the data. Sleep, 24, 165-170.
Bergen, B. (2012). Louder than words: The new science of how the mind makes meaning. New York: Basic Books.
Bischof, M., & Basset, C. (2004). Total dream loss: A distinct neuropsychological dysfunction after bilateral PCA stroke. Annals of Neurology, 56, 583-586.
Blake, Y., Terburg, D., Balchin, R., van Honk, J., & Solms, M. (2019). The role of the basolateral amygdala in dreaming. Cortex, 113, 169-183. doi:10.1016/j.cortex.2018.12.016
Braun, A., Balkin, T., Wesensten, N., Carson, R., Varga, M., Baldwin, P., Selbie, S., & Herscovitch, P. (1997). Regional cerebral blood flow throughout the sleep-wake cycle: An (H2O)-O-15 PET study. Brain, 120, 1173-1197.
Braun, A., Balkin, T., Wesensten, N., Gwadry, F., Carson, R., Varga, M., Baldwin, P., Belenky, G., & Herscovitch, P. (1998). Dissociated pattern of activity in visual cortices and their projections during human rapid eye movement sleep. Science, 279, 91-95.
Burgess, P. W., Dumontheil, I., & Gilbert, S. J. (2007). The gateway hypothesis of rostral prefrontal cortex (area 10) function. Trends in Cognitive Sciences, 11. doi:10.1016/j.tics.2007.05.004
Calvo-Merino, B., Jones, A., Haggard, P., & Bettina, F. (2017). Embodiment and expertise effects on aesthetics judgments. Paper presented at the Meetings of the Society for Cognitive Neuroscience.
Chang, C., Liu, Z., Chen, M., Liu, X., & Duyn, J. (2013). EEG correlates of time-varying BOLD functional connectivity. Neuroimage, 72, 227-236.
Chow, H. M., Horovitz, S., Picchioni, D., Balkin, T., & Braun, A. (2013). Rhythmic alternating patterns of brain activity distinguish rapid eye movement sleep from other states of consciousness. Proceedings of the National Academy of Sciences, 110, 10300-10305.
Christoff, K. (2014). Thinking. In K. Ochsner & S. Kosslyn (Eds.), The Oxford Handbook of Cognitive Neuroscience (pp. 318-333). New York: Oxford University Press.
Christoff, K., & Gabrieli, J. D. (2000). The frontopolar cortex and human cognition: Evidence for a rostrocaudal hierarchical organization within the human prefrontal cortex. Psychobiology, 28, 168-186.
Christoff, K., Gordon, A., Smallwood, J., Smith, R., & Schooler, J. (2009). Experience sampling during fMRI reveals default network and executive system contributions to mind wandering. Proceedings of the National Academy of Sciences of the United States of America, 106, 8719-8724.
Christoff, K., Irving, Z., Fox, K., Spreng, R. N., & Andrews-Hanna, J. (2016). Mind-wandering as spontaneous thought: a dynamic framework. Nature Reviews Neuroscience, 17, 718-731.
Christoff, K., Prabhakaran, V., Dorfman, J., Zhao, Z., Kroger, J. K., Holyoak, K., & Gabrieli, J. D. (2001). Rostrolateral Prefrontal Cortex Involvement in Relational Integration during Reasoning. Neuroimage, 14, 1136-1149. doi:10.1006/nimg.2001.0922
Christoff, K., Ream, J. M., Geddes, L. P., & Gabrieli, J. D. (2003). Evaluating Self-Generated Information: Anterior Prefrontal Contributions to Human Cognition Behavioral Neuroscience, 117, 1161-1168. doi:10.1037/0735-7044.117.6.1161
Coughlin, C. (2016). Development of Episodic Prospection: Factors Underlying Improvements in Middle and Late Childhood. (Ph.D.), University of California, Davis, Davis, CA.
Coughlin, C., Robins, R., & Ghetti, S. (2019). Development of episodic prospection: Factors underlying improvements in middle and late childhood. Child Development, 90, 1109-1122 doi:10.1111/cdev.13001
D'Argembeau, A. (2020). Imagination and Self-Referential Thinking. In A. Abraham (Ed.), The Cambridge Handbook of the Imagination (pp. 354-372 ). New York: Cambridge University Press.
Dang-Vu, T., Schabus, M., Desseilles, M., Schwartz, S., & Maquet, P. (2007). Neuroimaging of REM sleep and dreaming. In D. Barrett & P. McNamara (Eds.), The new science of dreaming: Biological aspects (Vol. 1, pp. 95-113). Westport, CT: Praeger/Greenwood.
Dixon, M. L., Fox, K., & Christoff, K. (2014a). Evidence for rostro-caudal functional organization in multiple brain areas related to goal-directed behavior. Brain Research, 1572, 26-39. doi:10.1016/j.brainres.2014.05.012
Dixon, M. L., Fox, K., & Christoff, K. (2014b). A framework for understanding the relationship between externally and internally directed cognition. Neuropsychologia, 62, 321-330.
Dixon, M. L., Moodie, C. A., Goldin, P. R., Farb, N., Heimberg, R. G., Zhang, J., & Gross, J. J. (2022). Frontoparietal and Default Mode Network Contributions to Self-Referential Processing in Social Anxiety Disorder. Cognitive, Affective, & Behavioral Neuroscience, 22, 187-198.
Dixon, M. L., Vega, A., Andrews-Hanna, J., Mills, C., Spreng, R. N., Cole, M. W., & Christoff, K. (2018). Heterogeneity within the frontoparietal control network and its relationship to the default and dorsal attention networks. Proceedings of the National Academy of Sciences of the United States of America, 115, E1598-E1607.
Domhoff, G. W. (1996). Finding meaning in dreams: A quantitative approach. New York: Plenum.
Domhoff, G. W. (2011). The neural substrate for dreaming: Is it a subsystem of the default network? Consciousness and Cognition, 20, 1163-1174.
Domhoff, G. W. (2018). The emergence of dreaming: Mind-wandering, embodied simulation, and the default network. New York: Oxford University Press.
Domhoff, G. W. (2020a). Dreaming is imagination roaming freely, based on embodied simulation, and subserved by an unconstrained default network. In A. Abraham (Ed.), The Cambridge Handbook of the Imagination (pp. 676-691). New York: Cambridge University Press
Domhoff, G. W. (2020b). Four assertions i did not make about dreams: Reply to Mageo (2019). Dreaming, 30, 100-105. doi:10.1037/drm0000125
Domhoff, G. W. (2022). The neurocognitive theory of dreaming: The where, how, when, what, and why of dreams. Cambridge: MIT Press.
Domhoff, G. W., & Fox, K. (2015). Dreaming and the default network: A review, synthesis, and counterintuitive research proposal. Consciousness and Cognition, 33, 342-353.
Dresler, M., Wehrle, R., Spoormaker, V., Koch, S., Holsboer, F., Steiger, A., ... Czisch, M. (2012). Neural correlates of dream lucidity obtained from contrasting lucid versus non-lucid REM sleep: A combined EEG/fMRI case study. Sleep, 35, 1017-1020.
Eichenlaub, J. B., Bertrand, O., & Ruby, P. (2014). Brain reactivity differentiates subjects with high and low dream recall frequencies during both sleep and wakefulness. Cerebral Cortex, 24, 1206-1215. doi:10.1093/cercor/bhs388
Eisbach, A. O. (2013). "Catching" a Wandering Mind: Developmental Changes in the Reporting of Off-Task Thoughts. Paper presented at the Society for Research on Child Development, Seattle.
Fair, D., Cohen, A. L., Dosenbach, N., Church, J. A., Miezin, F. M., Barch, D. M., ... Schlaggar, B. L. (2008). The maturing architecture of the brain's default network. Proceedings of the National Academy of Sciences, 105, 4028-4032.
Fair, D., Cohen, A. L., Power, J., Dosenbach, N., Church, J. A., Miezin, F. M., ... Peterson, S. (2009). Functional Brain Networks Develop from a ''Local to Distributed'' Organization. PLoS Biology, 5(doi:10.1371/journal.pcbi.1000381), 1-14.
Fosse, R., Hobson, J. A., & Stickgold, R. (2003). Dreaming and episodic memory: A functional dissociation? Journal of Cognitive Neuroscience, 15, 1-9.
Foulkes, D. (1982). Children's dreams: Longitudinal Studies. New York: Wiley.
Foulkes, D. (1999). Children's dreaming and the development of consciousness. Cambridge, MA: Harvard University Press.
Foulkes, D. (2017). Dreaming, reflective consciousness, and feelings in the preschool child. Dreaming, 27, 1-13.
Foulkes, D., Hollifield, M., Sullivan, B., Bradley, L., & Terry, R. (1990). REM dreaming and cognitive skills at ages 5-8: A cross-sectional study. International Journal of Behavioral Development, 13, 447-465.
Fox, K., Andrews-Hanna, J., & Christoff, K. (2016). The neurobiology of self-generated thought from cells to systems: Integrating evidence from lesion studies, human intracranial electrophysiology, neurochemistry, and neuroendocrinology. Neuroscience, 335, 134-150.
Fox, K., Nijeboer, S., Solomonova, E., Domhoff, G. W., & Christoff, K. (2013). Dreaming as mind wandering: Evidence from functional neuroimaging and first-person content reports Frontiers in Human Neuroscience, 7(Article 412), 1-18.
Fox, K., Spreng, R. N., Ellamil, M., Andrews-Hanna, J., & Christoff, K. (2015). The wandering brain: Meta-analysis of functional neuroimaging studies of mind-wandering and related spontaneous thought processes. Neuroimage, 111, 611-621.
Gackenbach, J. (1988). The psychological content of lucid versus nonlucid dreams. In J. Gackenbach & S. LaBerge (Eds.), Conscious mind, sleeping brain: Perspectives on lucid dreaming (pp. 181-220). New York: Plenum.
Gibbs, R. (2006). Embodiment and cognitive science. New York: Cambridge University Press.
Gopnik, A. (2009). The philosophical baby: What children's minds tell us about truth, love, and the meaning of life. New YorkFarar, Straus, and Giroux.
He, W., Sowman, P., Brock, J., Etchell, A., Stam, C., & al., e. (2019). Increased segregation of functional networks in developing brains. Neuroimage, 200, 607-620. doi:10.1016/j.neuroimage.2019.06.055
Henseler, I., Krüger, S., Dechent, P., & Gruber, O. (2011). A gateway system in rostral PFC? Evidence from biasing attention to perceptual information and internal representations Neuroimage, 56, 1666-1676. doi:10.1016/j.neuroimage.2011.02.056
Jus, A., Jus, K., Villeneuve, A., Pires, A., Lachance, R., Fortier, J., & Villeneuve, R. (1973). Studies on dream recall in chronic schizophrenic patients after prefrontal lobotomy. Biological Psychiatry, 6, 275-293.
Kerr, N., Foulkes, D., & Jurkovic, G. (1978). Reported absence of visual dream imagery in a normally sighted subject with Turner's syndrome. Journal of Mental Imagery, 2, 247-264.
Landau, M., Meier, B., & Keefer, L. (2010). A metaphor-enriched social cognition. Psychological Bulletin, 136, 1045-1067. doi:10.1037/a0020970
LeDoux, J. (2015). Anxious. New York: Viking.
LeDoux, J. (2019). The Deep History of Ourselves: The Four-Billion Year Story of How We Got Conscious Brains. New York: Viking.
LeDoux, J. (2020). How Does the Non-Conscious Become Conscious? Current Biology, 30, R1-R4. doi:10.1016/j.cub.2020.01.033.
LeDoux, J. (2021). What emotions might be like in other animals. Current Biology, 31, R824-R829.
LeDoux, J., & Lau, H. (2020). Seeing consciousness through the lens of memory. Current Biology, 30, R1018-R1022. doi:10.1016/j.cub.2020.08.008
LeDoux, J., Michel, M., & Lau, H. (2020). A little history goes a long way toward understanding why we study consciousness the way we do today. Proceedings of the National Academy of Sciences, 117, 6976-6984.
Lucidity Newsletter. (1986). A panel discussion of lucid dreaming. Lucidity Newsletter, 74-95.
MacDonald, S., & Culham, J. (2015). Do human brain areas involved in visuomotor actions show a preference for real tools over visually similar non-tools? Neuropsychologia, 77, 35-41.
Malinowski, J., & Horton, C. (2014). Memory sources of dreams: The incorporation of autobiographical rather than episodic experiences. Journal of Sleep Research, 23 441-447. doi:10.1111/jsr.12134
Mandler, J. (2008). On the birth and growth of concepts. Philosophical Psychology, 21, 207-230.
Mandler, J. (2012). On the spatial foundations of the conceptual system and its enrichment. Cognitive Science, 36, 421-451.
Maquet, P., Peters, J., Aerts, J., Delfiore, G., Dequerldre, C., Luxen, A., & Franck, G. (1996). Functional neuroanatomy of human rapid-eye-movement sleep and dreaming. Nature, 383, 163-166.
Mason, R., & Just, M. A. (2020). Neural Representations of Procedural Knowledge. Psychological Science, 31, 729-740.
McCaig, R. G., Dixon, M. L., Keramatian, K., Liua, I., & Christoff, K. (2011). Improved modulation of rostrolateral prefrontal cortex using real-time fMRI training and meta-cognitive awareness. Neuroimage, 55, 1298-1305.
Meyer, S., & Shore, C. (2001). Children's understanding of dreams as mental states. Dreaming, 11, 179-194.
Moraczewski, D., Nketia, J., & Redcay, E. (2020). Cortical temporal hierarchy is immature in middle childhood. Neuroimage, 216, 116616. doi:10.1016/j.neuroimage.2020.116616
Nelson, K. (2004). A welcome turn to meaning in infant development: Commentary on Mandler's The foundations of mind: Origins of conceptual thought. Developmental Science, 7, 506-507.
Nelson, K. (2005). Emerging Levels of Consciousness in Early Human Development. In H. S. Terrace & J. Metcalfe (Eds.), The missing link in cognition: Origins of self-reflective consciousness (pp. 116-141). New York: Oxford University Press.
Nelson, K. (2007). Young minds in social worlds: Experience, meaning, and memory. Cambridge: Harvard University Press.
Nelson, K., & Fivush, R. (2020). The development of autobiographical memory, autobiographical narratives, and autobiographical consciousness Psychological Reports, 123, 71-96. doi:10.1177/0033294119852574
Nofzinger, E., Mintun, M., Wiseman, M., Kupfer, D., & Moore, R. (1997). Forebrain activation in REM sleep: An FDG PET study. Brain Research, 770, 192-201.
Picchioni, D., Pixa, M. L., Fukunaga, M., Carr, W., Horovitz, S., Braun, A. R., & Duyn, J. H. (2014). Decreased connectivity between the thalamus and the neocortex during human nonrapid eye movement sleep. Sleep, 37, 387-397.
Poza, J., & Marti-Masso, J. (2006). Total dream loss secondary to left temporo-occipital brain injury. Neurologia, 21, 152-154.
Rolston, D. (2013). The Dream Catcher: Why does the researcher who pioneered the study of lucid dreaming face a future in academic exile? The Matter Archive Online, https://medium.com/matter-archive/the-dream-catcher-c85e83bb29693.
Sämann, P., Wehrle, R., Hoehn, D., Spoormaker, V., Peters, H., Tully, C., ... Czisch, M. (2011). Development of the brain's default mode network from wakefulness to slow wave sleep. Cerebral Cortex, 21, 2082-2093.
Schacter, D., Addis, D., & Buckner, R. (2008). Episodic simulation of future events: Concepts, data, and applications. Annals of the New York Academy of Sciences, 1124, 39-60. doi:10.1196/annals.1440.001
Sherman, L., Rudie, J., Pfeifer, J., Masten, C., McNealy, K., & Dapretto, M. (2014). Development of the default mode and central executive networks across early adolescence: A longitudinal study. Developmental Cognitive Neuroscience, 10, 148-159.
Solms, M. (1997). The neuropsychology of dreams: A clinico-anatomical study. Hillsdale, NJ: Erlbaum.
Spreng, R. N., Madore, K., & Schacter, D. (2018). Better imagined: Neural correlates of the episodic simulation boost to prospective memory performance Neuropsychologia, 113, 22-28. doi:10.1016/j.neuropsychologia.2018.03.025
Stevner, A., Vidaurre, D., Cabral, J., Rapuano, K., Nielsen, S., Tagliazucchi, E., ... Kringelbach, M. (2019). Discovery of key whole-brain transitions and dynamics during human wakefulness and non-REM sleep Nature Communications, 10(1035), 1-14. doi:10.1038/s41467-019-08934-3
Strauch, I. (2004). Traume im Ubergang von der Kindheit ins Jugendalter: Ergebnisse einer Langzeitstudie (Dreams in the Transition from Childhood to Adolescence: Results From a Longitudinal Study). Bern: Huber.
Strauch, I. (2005). REM dreaming in the transition from late childhood to adolescence: A longitudinal study. Dreaming, 15, 155-169.
Strauch, I., & Lederbogen, S. (1999). The home dreams and waking fantasies of boys and girls ages 9-15. Dreaming, 9, 153-161.
Tagliazucchi, E., von Wegner, F., Morzelewski, A., Brodbeck, V., Jahnke, K., & Laufs, H. (2013). Breakdown of long-range temporal dependence in default mode and attention networks during deep sleep Proceedings of the National Academy of Science, 110, 15419-15424.
Tarun, A., Wainstein-Andriano, D., Perogamvros, L., Solms, M., Schwartz, S., & Van De Ville, D. (2021). NREM sleep stages specifically alter dynamical integration of large-scale brain network. iScience, 24. doi:10.1016/j.isci.2020.101923
Taylor, M. (2013). Imagination. In P. D. Zelazo (Ed.), The Oxford handbook of developmental psychology (Vol 1): Body and mind (pp. 793-831). New York: Oxford University Press.
Tulving, E. (2005). Episodic memory and autonoesis. In H. S. Terrace & J. Metcalf (Eds.), The missing link in cognition: Origins of self-reflective consciousness (pp. 3-56). New York: Ocford University Press.
Uitermarkt, B. D., Bruss, J., Hwang, K., & Boes, A. D. (2020). Rapid eye movement sleep patterns of brain activation and deactivation occur within unique functional networks. Human Brain Mapping(June), 1-9. doi:10.1002/hbm.25102
Voss, U., Holzmann, R., Tuin, I., & Hobson, J. A. (2009). Lucid dreaming: A state of consciousness with features of both waking and non-lucid dreaming. Sleep, 32, 1191-1200.
Wilson, M. (2002). Six views of embodied cognition. Psychonomic Bulletin & Review, 9, 625-636.
Windt, J. (2015). Dreaming: A conceptual framework for philosophy of mind and empirical research. Cambridge: MIT Press.
Woolley, J. D., & Boerger, E. (2002). Development of beliefs about the origins and controllability of dreams Developmental Psychology, 38, 24-41.
Woolley, J. D., & Wellman, H. M. (1992). Children's conceptions of dreams. Cognitive Development, 7(3), 365-380.
 Go back to the Dream Library index. Go back to the Dream Library index.
|